Questo articolo e’ stato scritto, in occasione del Darwin day 2013, per il Carnevale della Biodiversita’ – ottava edizione, dal tema: “L’isola che c’è”. Per la rassegna completa di tutti i blog e post che partecipano al Carnevale vai sul blog Leucophaea, di Marco Ferrari.

Domenica, 7 ottobre 1492
[…] In questo giorno al levar del sole, la caravella Niña, che precedeva le altre perché più spedita, e tutte andavano a gara per vedere terra per primi […]
Giovedì, 11 ottobre 1492
[…] Videro gabbiani e un giunco verde vicino alla nave. Quelli della caravella Pinta scorsero un altro piccolo tronco, intagliato a quanto sembrava con ferro, e un pezzo di altra canna e altra erba, di quella di terra e una piccola tavola. Quelli della caravella Niña videro anche altri segnali di terra e un piccolo ramoscello carico di rose canine. Visti che ebbero questi segnali, tutti si rincuorarono e andarono lieti. […]
Venerdì, 12 ottobre 1492
[…] Alle due, passata la mezzanotte, apparve terra […] Giunti a terra, videro alberi verdissimi, molte fonti e frutti di varie sorte. […]
E’ così che Cristoforo Colombo raccontava, nel suo diario di bordo, l’approdo alle sue agognate Indie, che però non erano “altro” che S. Salvador e isole annesse (Cuba e Haiti). Un errore di poche migliaia di chilometri ma giustificabile per le conoscenze dell’epoca, che segnò l’arrivo dell’uomo occidentale in America. Nonostante non fosse un botanico, Colombo era un grande osservatore e quindi notò immediatamente la ricchezza e la diversità delle nuove terre, sia in senso di biodiversità che di diversità rispetto alle piante che conosceva, e ne faceva riferimento spesso nelle pagine del suo diario.
Non soltanto Colombo, ma anche molti altri navigatori di quel periodo facevano ben attenzione agli uccelli e ai vegetali che galleggiavano in acqua. In condizioni di scarsità di provviste e di acqua, con ciurme malate ed esasperate, spesso veri e propri covi di criminali, prive di medicinali e di donne, l’avvistamento della terra era di primaria importanza, se non altro per la vita stessa dell’ammiraglio e per le eventuali ricompense di regnanti finanziatori seduti sulle comode poltrone dei loro regni. E così, l’occhio del navigante era ben allenato a individuare subito uccelli di terra ma che di giorno andavano in mare per cibarsi, come gabbiani e cormorani, e a discernere una semplice erbetta galleggiante di alghe morte da residui di piante provenienti dalla costa. Questi erano infatti chiari segnali del vicino approdo. Ed è qui che entriamo in scena noi. Perché tra questi residui galleggianti ci sono i più grandi semi del mondo: le noci di cocco. I primi esploratori spagnoli le chiamarono “coco”, che significa “faccia di scimmia“, perché le tre tacche (occhi) sul dado peloso assomigliano un po’ alla testa e alla faccia pelosa di una scimmia.
La noce di cocco più comune è il seme della palma da cocco (Cocos nucifera), ma ce n’è anche un’altra un po’ meno conosciuta che è il seme della Lodoicea maldivica seychellarum, il cui trinomio topografico mi instilla inspiegabilmente un’incredibile invidia. Il seme di quest’ultima palma è il più grande e pesante in assoluto, ha una forma bifida e ricorda il bacino di una donna (da cui anche l’altro bel nome di Lodoicea callypige, cioè “dalle belle natiche”, di rimembranza ellenistica). Prima del 1800 si pensava che i semi giungessero dagli abissi marini e che si trattasse dei frutti di un albero che cresceva sotto la superficie del mare, conosciuto anche in Occidente come “cocco di mare” (“coco de mer” in francese). L’unico problema è che il seme maturo di Lodoicea non ha un mallo molto sviluppato (il mallo è il mesocarpo polposo tipico di una drupa, presente anche nelle nostre noci), ha un peso specifico maggiore di quello dell’acqua marina, e quindi non galleggia molto e non è trasportato dalle correnti marine se non per poche centinaia di metri. Per cui, dimentichiamocelo per ora.
La noce di cocco “classica” (quella, per intenderci, di Cocos nucifera, di cui da ora in poi parleremo) ha invece di un mallo fibroso molto leggero, resistente e ricco di aria che avvolge la noce, permettendo il suo galleggiamento per periodi di tempo molto lunghi e il trasporto mediante le correnti acquatiche (idrocoria). L’acqua marina, che distrugge la maggior parte degli altri semi, impiega molto tempo prima di penetrare nel mallo e nel guscio legnoso delle noci di cocco, le quali possono tranquillamente resistere fino a tre mesi in mare, a volte trasportate per centinaia di chilometri. Superato il punto di risacca delle spiagge di altre isole, i simpatici semini, sotto l’acqua piovana, germinano. E, dopo alcuni anni, il ciclo lentamente ricomincia. I frutti sono talmente duri e pesanti (fino a 2 chili e 30 cm di altezza) che ogni anno fanno fuori circa 1000 incauti turisti (non oso pensare ai possibili epigrammi), molti più delle vittime degli squali ad esempio.


Noci di cocco (in alto) di Lodoicea callypige e (in basso) di Cocos nucifera.
Come negli animali, anche tra le piante ci sono specie a strategia r e specie a strategia K . Le prime sono di solito annuali e monocarpiche adattate a riprodursi rapidamente e a colonizzare habitat nuovi e spesso transitori, dove la competizione è bassa. Le seconde hanno una vita più lunga, sono spesso perenni e policarpiche, e conservano una parte della loro energia per la loro crescita vegetativa. Nel nostro caso, con frutti grandi e numerosi, ci troviamo di fronte ad una specie a strategia K, per nulla affatto pioniera, in grado di crescere velocemente come una palma sa fare, ma solo in determinate condizioni di luce, acqua e temperatura; quindi abbastanza esigente.
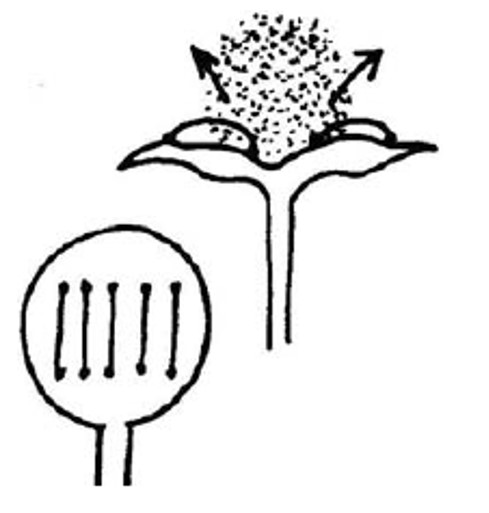
Molte specie vegetali sono in grado di disperdersi per mezzo dell’acqua. E’ noto che la vita terrestre, e questo vale anche per le piante, derivi dall’acqua. Per questo, le prime piante eterosporee (con gameti “maschili” e “femminili” diversi in forma, dimensione e disposizione) avevano megaspore femminili che si disperdevano efficientemente attraverso l’acqua. Le felci che vivevano nelle paludi del Carbonifero (350 milioni di anni fa) si propagavano presumibilmente per idrocoria. Ad esempio, Lepidocarpon , una felce arborea che raggiugeva dimensioni imponenti (fino a 40 m di altezza), estintasi alla fine del Carbonifero, aveva una megaspora che funzionava prima da paracadute e poi, una volta in acqua, era trasportata dal vento come una vela (e la vela era un organo dal leggiadro e semplice nome di “lepidostrobofillo”).

Megasporangio di Lepidocarpon. Fonte: http://www.ucmp.berkeley.edu/IB181/VPL/Lyco/Lyco3.html
I discendenti tuttora viventi di queste felci arboree, appartenenti al genere Isoetales, presentano poche specie che vivono in zone umide con ruscellamento. Ancora oggi, molte specie di felci sono prevalentemente acquatiche o perlomeno idrofile. Molti sono i generi di piante acquatiche che si riproducono vegetativamente per frammentazione, quali Elodea, Hydrilla e Lagariosiphon, tristemente diventate “erbacce acquatiche” dopo la loro introduzione in nuovi ambienti da parte dell’uomo, ma l’idrocoria, cioè la dispersione di semi e frutti per mezzo dell’acqua, è tipica di poche specie.
I frutti e i semi dispersi in acqua sono relativamente grandi, e a volte molto grandi. Le noci di cocco siamesi di Lodoicea maldivica pesano tutte e due sui 15-30 chili e possono arrivare a mezzo metro di lunghezza, conquistando così il primato di frutto (e seme) più grande del mondo! Le più ordinarie noci di cocco di Cocos nucifera hanno un seme soltanto, ma comunque bello grosso. I frutti sono infatti drupe ovali voluminose, con tre spigoli arrotondati e sono provvisti di tre involucri: il più esterno è un’epidermide (epicarpo) liscia e sottile, inizialmente di colore verde, che a maturazione diventa prima giallastra e poi bruna; al di sotto di essa vi è lo spesso strato fibroso (mesocarpo) che racchiude al suo interno un endocarpo legnoso durissimo; nelle noci immature esso racchiude a sua volta l’endosperma oleoso (tessuto ricco di nutrienti destinato ad alimentare l’embrione), o mandorla – in pratica, quello tanto reclamizzato ad alta voce sulle bancarelle – cavo all’interno e ripieno di un succo opalescente lattiginoso, di sapore fresco e zuccherino (“succo di cocco”), che diminuisce in quantità man mano che il frutto matura e che l’italiano medio fa fuoriuscire infilzando con delicatezza un cacciavite in uno dei tre fori con l’aiuto di un martello. L’embrione è situato all’interno ad un’estremità della mandorla. I tre strati nel loro insieme costituiscono il frutto; l’endosperma e l’embrione rappresentano invece il seme. L’involucro legnoso durissimo presenta alla base tre pori dalla superficie morbida, detti occhi: è da uno di essi che il germoglio originatosi dall’embrione avrà la possibilità, perforandolo, di fuoriuscire dal guscio, dando così origine alla nuova plantula.
Anche da sola, la noce di cocco è contemporaneamente una bevanda, un alimento e una fibra. Fornisce acqua, latte e olio per cucinare. Il chiaro e dolciastro succo di cocco è una bevanda rinfrescante. Dal momento che questo fa parte dell’endosperma, il quale nutre l’embrione, il succo è ricco di fitormoni per la crescita (le citochinine e altri composti simili sono stati identificati per la prima volta proprio dal succo di cocco, e questo è spesso addizionato ai terreni di crescita usati nelle colture cellulari vegetali e nella micropropagazione). Il latte di cocco, un po’ come il nostrano di mandorla, si ottiene invece mescolando la polpa grattugiata con acqua e poi spremendo il tutto per far uscire il liquido; esso insaporisce e arricchisce zuppe, salse e impasti. Per estrarre l’olio dalle noci di cocco, usato direttamente per cucinare nei paesi tropicali e per preparare altri prodotti nei paesi occidentali, si deve spaccare la noce matura. Dopo essiccazione al sole, la polpa, detta copra, viene separata dal guscio per estrarne l’olio. Infine, le fibre dell’endocarpo legnoso, costituite da piccoli fili lignificati lunghi circa a 1 mm, una volta lavorate, raggiungono anche i 30 cm di lunghezza; sono leggere, elastiche e resistenti all’abrasione, e vengono utilizzate per fabbricare pennelli, scope e cordame.

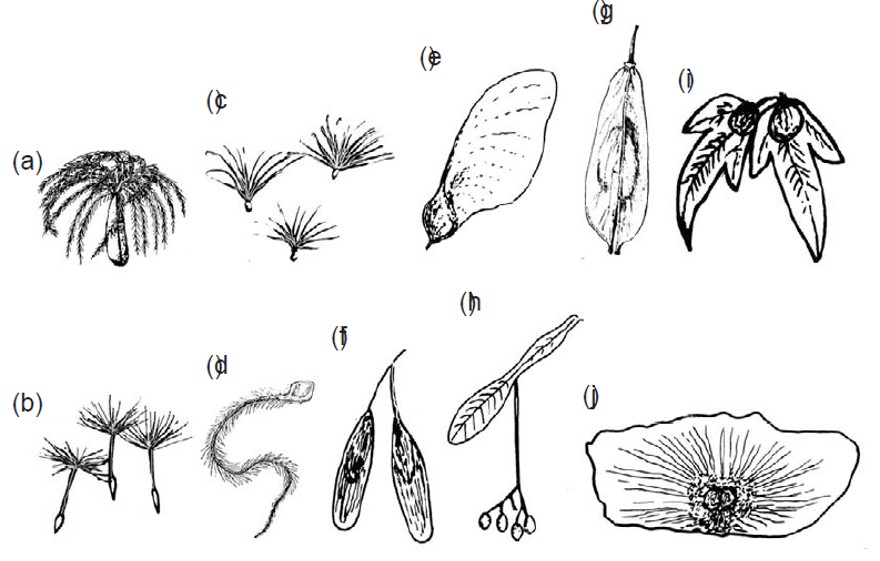
Alcune specie idrocore: (a) Lodoicea maldivica, (b) Cocos nucifera, (c) Entada gigas. Fonte: Ingrouille e Eddie (2006).
Oltre alle noci di cocco, un altro caso di gigantismo è quello di Entada gigas, una pianta rampicante legnosa che produce legumi di 12 centimetri di larghezza e più di 1 m di lunghezza. Il frutto si rompe in segmenti fluttuanti, ciascuno contenente un grosso seme, che si riversano nei corsi d’acqua. Si suppone sia stata l’osservazione dei semi di Entada sulle spiagge delle Azzorre che abbia spinto Cristoforo Colombo a ipotizzare l’esistenza di un continente di là dall’Atlntico. Ancora oggi, gli inglesi chiamano quasi semi “seabens” perché arrivano sulle loro coste trasportati dalla corrente del Golfo.
Alcuni semi e frutti di piante idrocore galleggiano grazie alle della presenza di camere d’aria, come nel frutto di Xanthium o nel falso frutto di Atriplex. Il frutto-scatola (Barringtonia asiatica) del sud-est asiatico può rimanere a galla per almeno due anni ed è anche utilizzato come galleggiante per la pesca.


(In alto) Falso frutto di Atriplex halimus, munito di camere d’aria. Bari, 1 m, ott 2012. Foto di Vito Buono. (In basso) Frutto-scatola (Barringtonia asiatica). Foto di Barry Conn e Kipiro Damas (http://www.pngplants.org/PNGtrees/TreeDescriptions/Barringtonia_asiatica_L_Kurz.html).
Si stima che solo circa 250 specie sono regolarmente disperse nelle isole oceaniche mediante acqua ed è stato stimato che, dei 378 colonizzazioni vegetali originari delle isole Galapagos, solo il 9% sono idrocore (le restanti sono state trasportate dal vento o uccelli). Non è solo una questione di sopravvivenza durante il periodo di dispersione in mare. Stabilizzarsi nella zona intertidale, infatti, non è per nulla semplice. Per questo motivo, anche qui la selezione naturale si è scatenata: ad esempio, la mangrovia rossa (Rhizophora mangle) si aiuta con la viviparità, producendo una lunga piantina pendente prima che venga definitivamente rilasciata. Non si tratta di propaguli perché la riproduzione non è vegetativa, ma di vere e proprie piantine. Un meccanismo simile è presente nella Aegialitis (Plumbaginaceae). Quest’ultima è stata uno delle prime specie tropicali a raggiungere e Krakatoa dopo una catastrofica eruzione vulcanica avvenuta nell’agosto del 1883.


Giovani piantine pendenti (in alto) di mangrovia rossa (Rhizophora mangle) e (in basso) di Aegialitis.
Alcune piante acquatiche non sono strettamente idrocore ma ittiocore, cioè disperse dai pesci, o avicore, cioè diffuse da uccelli acquatici. I frutti dell’erba acquatica Glyceria, sono consumati da carpe; i frutti a forma di oliva della infestante acquatica Posidonia adottano la tattica “Pinocchio-style” e sono mangiati dai tonni nel Mediterraneo; e ancora, i peli sul frutto delle canne del genere Typha prevengono l’imbibizione prima della liberazione, ma in seguito permettono il galleggiamento in acqua, dove si aprono e affondano, per infine germinare (ma alcuni dei suoi semi appuntiti sono stati trovati aderenti alla pelle dei pesci).
I semi delle ninfee maturano nel frutto sott’acqua ma, quando rilasciati, riemergono a galla in massa, dove diventano appetibili per gli uccelli acquatici. Come per molti semi di piante terrestri, il passaggio attraverso l’intestino degli uccelli acquatici può agire come uno stimolante per la loro germinazione. In Nuphar, i frutti maturano sopra la superficie dell’acqua, ma sono poi i carpelli contenenti i semi che galleggiano come esche sulla superficie, dove sono mangiati dai pesci. Molti frutti delle piante acquatiche (alcune delle quali estinte, come Ceratophyllum e Trapa) sono duri e con escrescenze spinose, due caratteristiche che possono favorire la dormienza e la dispersione, nonché l’ancoraggio su substrati sono instabili. Le spine sui frutti della Victoria amazonica, le cui foglie galleggianti sono capaci di sorreggere un bambino, svolgono un ruolo diverso: quello della protezione dagli erbivori.


(In alto) Frutto-esca di Nuphar. (In basso) Victoria amazonica.
Anche senza citare i noti studi alle Galapagos di San Darwin, di cui oggi celebriamo il Natale (la mia teoria zichicca “scienza ≈ religione” prende sempre più piede), gli ecosistemi insulari sono sempre stati oggetto di particolare attenzione da parte di naturalisti ed ecologi. La discontinuità terra-acqua pone dei limiti ben precisi alla distribuzione delle specie rendendo le comunità insulari sostanzialmente chiuse ad interazioni ecologiche con l’esterno. La diversità nelle isole ha quindi delle caratteristiche molto interessanti; ed è per questo che sono state e sono da sempre molto studiate. L’immigrazione delle specie dalla terraferma all’isola diventa tanto più difficile quanto maggiore è la distanza dalla costa. Già nel 1967, MacArthur e Wilson avevano ipotizzato che il numero di specie presente su di un’isola variasse come risultato di due forze contrapposte. Da una parte, specie non ancora presenti sull’isola possono giungere sull’isola dalla terraferma (anche portate su un’isola da “zattere” naturali o artificiali), dall’altra le specie già presenti sull’isola possono estinguersi. Infatti, le popolazioni delle specie insulari sono in generale molto più piccole di quelle ospitate dalla terraferma e quindi soggette a una serie di problemi che le possono condurre più facilmente all’estinzione rispetto ai più fortunati cospecifici continentali. Ciò non è accaduto alla noce di cocco, che anzi si è diffusa enormemente in tutti i paesi tropicali e, almeno inizialmente, soprattutto nelle isole. Perché?
Di isola in isola per sua girovaga natura, la noce di cocco ha un areale genetico di origine pressoché sconosciuto. L’argomento è infatti ancora un mistero irrisolto, sia perché i frutti si disperdono per mezzo delle correnti, sia perché è stata diffusa dai popoli che colonizzarono le isole oceaniche.
L’elevato dinamismo migratorio della palma da cocco ha fatto sì che il suo luogo d’origine sia ancora oggi uno dei grandi enigmi insoluti della biologia vegetale. Infatti, le sue fasi di diffusione, le vie migratorie, l’età e il luogo d’origine della palma da cocco non sono ricavabili con sicurezza partendo dalla distribuzione attuale. Si ritiene che sia originaria dell’arcipelago indo-malesiano e che nell’antichità si sia diffusa per mare nelle vicine isole del Pacifico, prima che l’uomo la trasportasse per distanze ancora maggiori, probabilmente a ovest, fino al sud dell’India e nello Sri Lanka e ad est nelle Isole Samoa. Negli ultimi 250 anni gli studiosi hanno proposto però anche teorie diverse, che prevedono l’origine in America Centrale, Polinesia, Fiji, ed altre aree ancora.
Gli europei (portoghesi e spagnoli) scoprirono per la prima volta il cocco esplorando le coste occidentali dell’America centro meridionale e dal 1525 cominciarono a coltivarlo diffondendolo altrove. Successivamente, l’ibridazione, la selezione e la diffusione da parte dell’uomo, ha dato alla vasta gamma di varietà e di distribuzione pantropicale che vediamo oggi.
Torniamo un attimo indietro: quindi in America il cocco era in qualche modo arrivato?
L’attestazione precolombiana del cocco in America centrale proviene da analisi dei microsatelliti del DNA delle popolazioni di palma da cocco in America Centrale e nel Pacifico. Sembra che in America la palma da cocco sia stata portata da popolazioni umane provenienti dal sud-est asiatico (il trasporto via mare effettivamente era difficile, data la lontananza!), il cui vasellame è stato ritrovato anche in Ecuador. Le analisi dei microsatelliti hanno dimostrato che il cocco americano è geneticamente più vicino a quello filippino rispetto a quello delle più vicine isole della Polinesia, rafforzando così le evidenze archeologiche.

Vasi a forma di casa: manufatto di origine sud-est asiatica ritrovati in Ecuador. Fonte: Baudouin & Lebrun (2009).
Questo per sud-est asiatico e America. Passiamo ora alla migrazione verso ovest. Analisi con marcatori RFLP, hanno confermato che le popolazioni di palma da cocco del sud-est asiatico e delle isole del Pacifico hanno più polimorfismi e sono quindi geneticamente più diverse, mentre le palme di India, Sri Lanka e Africa occidentale hanno caratteristiche genetiche più omogenee. Questo potrebbe significare che le zone di origine siano le prime e poi, per mano dell’uomo o per idrocoria, la specie si sia diffusa verso l’ovest.
In tutto questo marasma migratorio, intervengono le varietà nane (la palma da cocco “wild type” giunge anche a 25-30 metri di altezza), arrivate fino in Africa, e probabilmente introdotte all’inizio del 1900 dall’Asia e dal Pacifico. Si sarebbe trattato di pochi mutanti, considerando la bassa variabilità genetica di queste popolazioni. Il marasma migratorio della palma da cocco diventa manicomio allo stato puro se si considerano anche gli spostamenti tettonici e le isole più o meno temporanee che hanno fatto da ponte per la dispersione naturale di questa specie (se ne avete la forza, leggete il lavoro di Harries del 1990, sotto in bibliografia). Mentre il cocco viaggiava, infatti, i continenti si spostavano. Harries è però fortunatamente (per me) in accordo con altri botanici sul fatto che la palma da cocco sia stata coltivata per la prima volta proprio in Malesia, dove ha subito un vero programma empirico di miglioramento genetico da parte degli agricoltori locali.
Sfinito da cotanto girovagare non mi resta che consultare il lavoro di Gunn et al. (2011). L’articolo è recente (per molti revisori, segno incontrovertibile di garanzia). Qui, “nell’analisi genetica più ampia finora mai svolta” (umili, gli autori), sono state trovate prove di due distinte origini di coltivazione della palma di cocco: una nelle isole del sud-est asiatico, l’altra nei margini meridionali del subcontinente indiano. Nonostante il lungo e ampio movimento di palme dovuto agli uomini sia all’interno che tra questi bacini oceanici, le piante attuali non mostrano segni di sostanziale commistione genetica tra le due principali sottopopolazioni. Data l’assenza di evidenti barriere riproduttive, l’elevato livello di differenziazione genetica tra le due sottopopolazioni suggerisce un lungo periodo di isolamento prima dell’influenza umana. In questa luce, la predominanza di commistione genetica nella parte occidentale dell’Oceano Indiano suggerisce che gli esseri umani abbiano probabilmente giocato un ruolo di primo piano nella coltivazione e nella propagazione di palme di cocco in quella regione. Nel Pacifico, invece, la selezione umana ha determinato la comparsa di tratti utili quali il nanismo, la capacità di autoimpollinarsi e particolari forme del frutto. Infine, la sottopopolazione presente nel Madagascar è particolarmente varia, probabilmente a causa dell’antico percorso commerciale austronesiano, che connetteva est Africa, Madagascar, il sud-est asiatico, Formosa e Oceania. Gli Arabi contribuirono poi a diffondere la specie sulle coste dell’Africa con i loro commerci nell’Oceano Indiano.
Se avete ancora un po’ pazienza, vi lascio con questa mappa: è la versione più recente della distribuzione geografica delle sottopopolazioni di palma da cocco nel mondo secondo Gunn et al. (2011). La specie ne ha fatti di giri!

Distribuzione geografica delle popolazioni indo-atlantiche e pacifiche di palma da cocco secondo Gunn et al. (2011) (liberamente disponibile on-line).
Grazie a loro, ho scritto:
Baudouin L & Lebrun P (2009) Coconut (Cocos nucifera L.) DNA studies support the hypothesis of an ancient Austronesian migration from Southeast Asia to America. Genet Resour Crop Evol 56: 257-262.
Colombo C (1492-1493) Diario di Bordo. Cristoforo Colombo. Collana Storia d’Italia. Einaudi, Torino.
Dennis JV, Gunn CR (1971) Case against trans-Pacific dispersal of the coconut by ocean currents. Economic Botany 25(4): 407-413.
Diamond JM (1973) Distributional ecology of New Guinea birds. Science, 179:759-769.
Gunn BF, Baudouin L, Olsen KM (2011) Independent origins of cultivated coconut (Cocos nucifera L.) in the old world tropics. PLos ONE 6(6): e21143.
Harries HC (1990) Malesian origin for a domestic Cocos nucifera. In: The Plant Diversity of Malesia eds. P. Baas, K. Kalkman e R. Geesink. Pp. 351-357. Leyden.
Harries HC (1978) The evolution, dissemination and classification of Cocos nucifera L. The Botanical Review 44(3): 265-319.
Ingrouille M, Eddie B (2006) Plants: Evolution and Diversity. Cambridge University Press, UK.
Lebrun P, N’cho YP, Seguin M, Grivet L, Baudouin L (1998) Genetic diversity in coconut (Cocos nucifera L.) revealed by restriction fragment length polymorphism (RFLP) markers. Euphytica 101: 103-108.
MacArthur RH, Wilson EO (1967) The Theory of Island Biogeography. Princeton University Press, Princeton, NJ, USA.
Smith JMB (1991) Tropical drift disseminules on Southeast Australian beaches. Australian Geographical Studies 29(2): 355-369.
Tozzi M (2010) Ma fanno più vittime le noci di cocco. “La stampa” on-line.
Dalla Grande Ragnatela Mondiale:
http://it.wikipedia.org/wiki/Lodoicea_maldivica
http://it.wikipedia.org/wiki/Mallo_%28botanica%29
http://lastampa.it/2010/12/06/cultura/opinioni/editoriali/ma-fanno-piu-vittime-le-noci-di-cocco-vweaBadfG15FrKcZOgZaRM/pagina.html
http://olmo.elet.polimi.it/ecologia/dispensa/node68.html
http://wol.jw.org/it/wol/d/r6/lp-i/102003208
http://www.ucmp.berkeley.edu/IB181/VPL/Lyco/Lyco3.html

 Vischio (Viscum album L.) su un ramo di pero selvatico (foto: A. Sofo)
Vischio (Viscum album L.) su un ramo di pero selvatico (foto: A. Sofo) Pseudo-bacche di vischio (Viscum album L.) (foto: A. Sofo)
Pseudo-bacche di vischio (Viscum album L.) (foto: A. Sofo) Vischio tropicale; Loranthaceae (fonte: M. Ingrouille e W. Eddie 2006)
Vischio tropicale; Loranthaceae (fonte: M. Ingrouille e W. Eddie 2006) Mimetismo (riquadro in alto a destra) di Amyema linophylla
Mimetismo (riquadro in alto a destra) di Amyema linophylla Guano di tordela con semi di vischio (fonte: A. Girodo)
Guano di tordela con semi di vischio (fonte: A. Girodo)