Semi a propulsione
 Ecballium elaterium, Murgia di Spinazzola (BA). Foto mie.
Ecballium elaterium, Murgia di Spinazzola (BA). Foto mie.
Senza entrare nel merito della specie, la quale è stata abbondantemente studiata dal punto di vista botanico e farmaceutico, camminando nella sagra successiva all’escursione, tra castagne e cardoncelli, pensavo che le piante hanno sviluppato innumerevoli adattamenti nei loro semi, con ogni forma diversa sviluppata per adattarsi a un ambiente e un ciclo di vita unici. Da qui la curiosità di approfondire di più l’argomento, di solito poco affrontato nei corsi universitari (ricordo solo qualche accenno nei corsi di Botanica e Botanica Sistematica che ho frequentato un secolo fa).
La dispersione dei semi di una pianta è una fase estremamente importante del suo ciclo di vita. Questa capacità ha permesso alle piante di colonizzare nuove aree e di migrare in risposta alle mutevoli condizioni del clima e della competizione con altre piante, influenzando così sia la dinamica che la composizione delle comunità vegetali e animali. La produzione di semi è stata senza dubbio una delle maggiori innovazioni evolutive che si sono verificate durante la radiazione delle piante terrestri. Le specie vegetali hanno sviluppato tantissime varianti di semi, con ogni forma diversa sviluppata per adattarsi a un ambiente e un ciclo di vita unici. I primi produttori di semi, le Gimnosperme, comprendenti le estinte Pteridosperme, e le conifere e le cicadofite oggi esistenti, hanno “semi nudi”, cioè che non hanno tessuti (frutti) attorno. Quando i coni delle conifere (le pigne) si asciugano, le loro brattee simili a scaglie si aprono lentamente, rilasciando i semi, che cadono dall’albero fluttuando nella brezza usando strutture simili ad ali, per poi germinare lontano dalla pianta madre. Alcuni dei loro parenti, tra cui il ginkgo e il tasso, producono semi con rivestimenti carnosi colorati che attirano uccelli e mammiferi, i quali poi li disperdono.
Furono le Angiosperme, le piante con fiori, che videro alcune delle più grandi modifiche nel modo in cui le piante potevano diffondere il loro materiale genetico. Inizialmente comparvero semi soffici e leggeri tanto da farsi trasportare dal vento (anemocoria). Alcuni generi, come Asclepias, producono grandi baccelli dai quali numerosi minuscoli semi attaccati esplodono nell’aria, come sottili paracadute, mentre in Clematis (clematide) le “code” allungate dei semi permettono loro di essere trasportati anche dal più leggero zefiro. Altre specie, come aceri e sicomori, hanno evoluto semi a “pala di elicottero” che si allontanano trasportati dal vento e dalla gravità. In altre specie ancora, le strutture uniche dei semi possono essere rivelate solo a livello microscopico, come quelle del pennello indiano (Castilleja flava), che hanno una forma cava a nido d’ape in grado di catturare l’aria.
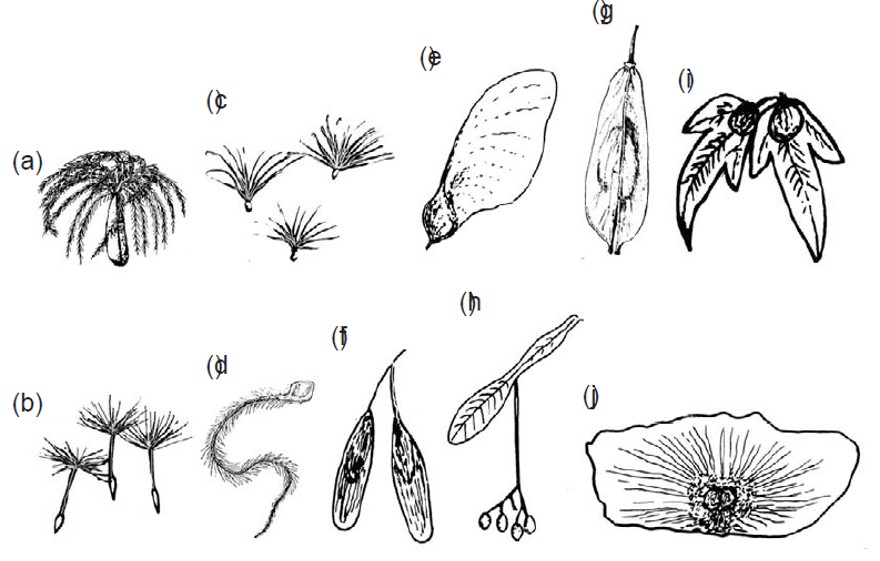 Esempi di anemocoria: semi e frutti alati o con “paracadute”: a) valeriana, b) tarassaco, c) Epilobium, d) Clematis, e) acero, f) frassino, g) Tipuana, h) tiglio, i) carpino, j) Alsomitra.
Esempi di anemocoria: semi e frutti alati o con “paracadute”: a) valeriana, b) tarassaco, c) Epilobium, d) Clematis, e) acero, f) frassino, g) Tipuana, h) tiglio, i) carpino, j) Alsomitra.
Tempo fa ho scritto anche un artiolo sull’idrocoria, cioè sul trasporto nell’acqua di semi, polline, spore e frutti, ma un’altra strategia che si è rivelata di grande successo, e qui torniamo al nostro Ecballium, è stata quella di produrre semi stretti saldamente in frutti che a maturità scoppiano. In questo caso rientriamo nell’autocoria, cioè nella disseminazione operata senza l’aiuto di agenti estranei alla pianta, detta anche autodisseminazione. Anche un piccolo mammifero che passa accanto ad una bacca matura di cocomero asinino causerà un’esplosione di semi fino a 12 metri di distanza e alla velocità di 10 m/s (pari a 36 km/h !); e un minimo colpo a un frutto del balsamina (Impatiens glandulifera) attiverà un’eruzione velocissima di semi scuri in tutte le direzioni. Altri esempi notevoli di autocoria sono l’eiaculatore delle Bignoniaceae, un funicolo modificato, e le capsule esplosive nel genere Viola. Alcune piante, come i papaveri, hanno frutti a forma di saliera che gettano i loro semi a terra mentre ondeggiano nel vento e urtano i loro vicini. Oppure, nel caso di molte orchidee, semi della dimensione della polvere possono essere trasportati e portati via da un unico soffio d’aria.
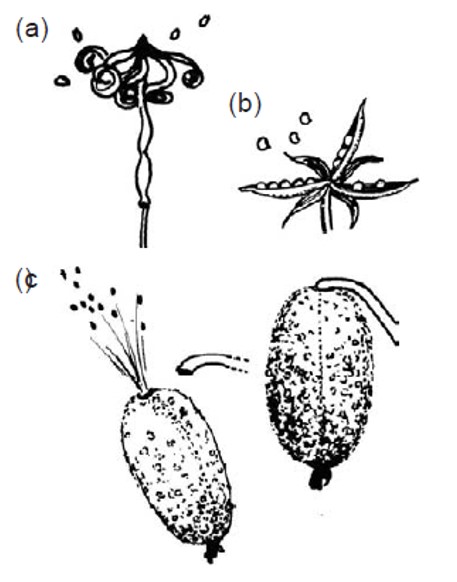 Esempi di autocoria: rilascio esplosivo di semi in a) Impatiens, b) viola e c) cocomero asinino.
Esempi di autocoria: rilascio esplosivo di semi in a) Impatiens, b) viola e c) cocomero asinino.
L’autocoria non è esclusiva dei semi, ma anche delle spore e del polline. La maggior parte delle piante adotta un rilascio passivo, per cui la posizione e la disposizione degli sporangi (dove maturano le spore) e delle antere (dove matura il polline) sopra o al di fuori di foglie, brattee o perianzio consentono al vento di trasportare le spore e il polline. Molte specie hanno un cono maschile, un amento o lunghi filamenti penzolanti da cui si distaccano le spore. La vibrazione di questi organi nella brezza può essere molto regolare ed è stato osservato che anche con un flusso d’aria moderato di 2-3 m/s, le infiorescenze di molte specie erbose si agitano in modo armonico.
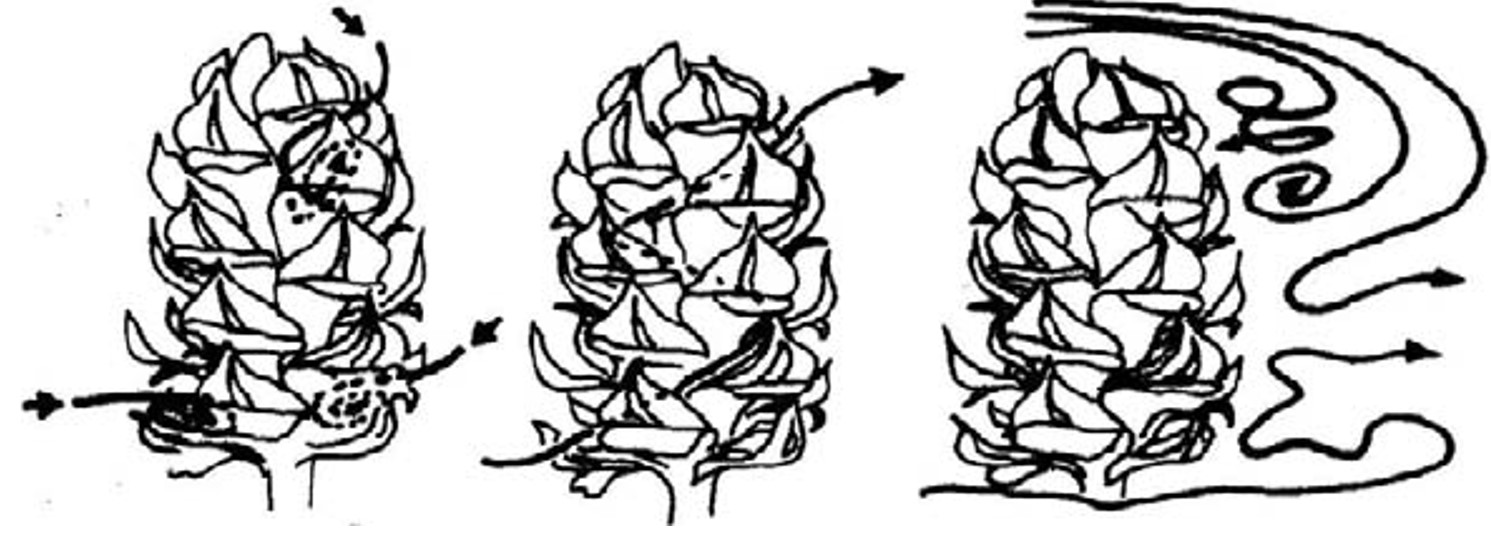 Cono femminile di pino dove è mostrato l’intrappolamento del polline trasportato dal vento.
Cono femminile di pino dove è mostrato l’intrappolamento del polline trasportato dal vento.
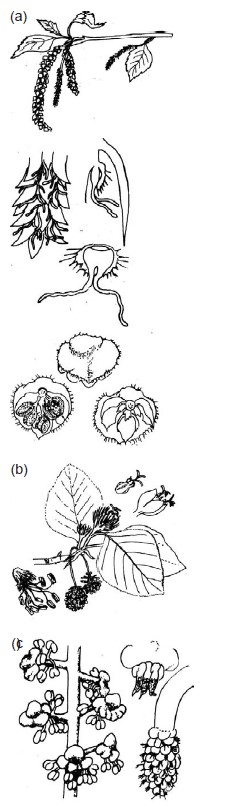 Amenti di Fagales ad impollinazione anemocora: fiori maschili e femminili di a) betulla, b) faggio e c) ontano.
Amenti di Fagales ad impollinazione anemocora: fiori maschili e femminili di a) betulla, b) faggio e c) ontano.
I meccanismi di rilascio delle spore sono anche alimentati da variazioni della pressione dell’acqua, che si verifica quando le celle si disidratano. A questo proposito, gli elateri sono cellule allungate con una o più bande di ispessimento che permettono loro ti torcersi quando l’umidità cambia, muovendosi quindi igroscopicamente all’interno della massa delle spore. Essi si trovano inframmezzati alle spore e ne favoriscono la dispersione con movimenti igroscopici. Gli elateri si trovano in tre gruppi di piante: epatiche, antocerote ed equiseti. In alcune specie di epatiche, gli elateri sono attaccati a un piccolo elatoforo colonnare, alla base (Pellia) o all’apice (Riccardia) della capsula. In Cephalozia, l’elaterio viene compresso e si attorciglia mentre si asciuga, fino a quando la colonna d’acqua al suo interno si rompe; quindi balza indietro, torcendosi per scrollarsi di dosso le spore. Gli elateri di Frullania sono attaccati alla parte superiore e inferiore delle valvole della capsula; quando le valvole si aprono, allungano gli elateri, che si strappano alla base rilanciando spore.
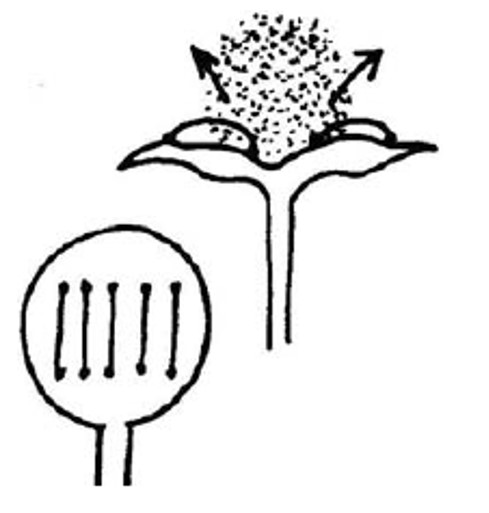 Azione degli elateri di Frullania: la capsula si apre dividendosi in lobi; gli elateri attaccati alla parete della capsula si allungano e poi rilasciano come una molla, catapultando le spore fuori.
Azione degli elateri di Frullania: la capsula si apre dividendosi in lobi; gli elateri attaccati alla parete della capsula si allungano e poi rilasciano come una molla, catapultando le spore fuori.
Di tutti i gruppi di piante senza semi, i muschi presentano la gamma più diversificata di meccanismi per la dispersione delle spore, con tantissime variazioni nella forma del peristoma. I denti del peristoma possono essere multicellulari e fatti di cellule intere o costruiti dalle parti di cellule adiacenti che si sono rotte a maturità. Il peristoma può essere attorcigliato e i denti possono essere biforcuti o filamentosi. Il restringimento della capsula di muschio spinge via l’opercolo a forma di tappo perché quest’ultimo si restringe meno del resto della capsula, spingendo così anche le spore verso l’apertura, dove i denti del peristoma, muovendosi igroscopicamente, sollevano le spore nel flusso d’aria. In alcuni generi, come Bryum, esiste un doppio peristoma: l’anello esterno si interdigita tra quello l’interno e raccoglie le spore sulle punte segmentate dentate, facendole sfuggire. Nei muschi di palude Sphagnum, l’opercolo esplode, sparando spore, a causa della pressione che si accumula all’interno quando la capsula si asciuga e si restringe.
 Azione della capsula di Sphagnum durante il rilascio delle spore. L’opercolo esplode, lanciando le spore fuori quando la capsula si restringe a maturità.
Azione della capsula di Sphagnum durante il rilascio delle spore. L’opercolo esplode, lanciando le spore fuori quando la capsula si restringe a maturità.
Nelle felci leptosporangiate, c’è uno speciale anello di cellule, l’annulus, che aiuta a catapultare le spore dallo sporangio; le cellule anulari si restringono mentre si disidratano e alla fine lo sporangio si rompe lungo lo stomio in due parti, di cui una recante le spore. Immediatamente, l’acqua liquida all’interno delle cellule dell’anello si vaporizza sotto la tensione, rilasciando così la pressione in modo che le due parti tornino l’una vicino all’altra e le spore vengano catapultate nell’aria. Nell’ortica (Urtica dioica) gli stami sono curvati verso l’interno a mo’ di bocciolo e i filamenti sono piegati all’indietro come molle. Con il tempo asciutto, i filamenti scattano all’indietro, lanciando polline nell’aria. Quando il bocciolo di Broussonettia si apre, i filamenti spuntano e i granelli di polline vengono lanciati violentemente in aria. Infine, la mercorella comune (Mercurialis annua) ha peduncoli che si rompono, lanciando i fiori e il loro polline nell’aria.
Ma le piante da fiore non si limitano a usare solo i mezzi abiotici di vento, acqua e propulsione a getto per diffondere i loro semi. Poiché le piante da fiore si sono gradualmente irradiate e si sono differenziate a partire dal Cretaceo (circa tra 145 e 65 milioni di anni fa), il ruolo degli animali è diventato sempre più importante per la loro sopravvivenza. I semi appiccicosi di Galium aparine si sono evoluti per aderire agli uccelli e ai mammiferi che sfiorano la sua vegetazione gommosa, e così anche i semi uncinati di Hackelia americana si sono adattati per avere un passaggio dagli animali di passaggio, aggrappandosi alla loro pelliccia. Questi metodi semplici ma efficaci di dispersione dei semi si sono rivelati molto vantaggiosi.
Di pari passo con l’evoluzione dei fiori, un gran numero di specie animali divenne presto necessario per trasportare polline, e col tempo anche le Angiosperme iniziarono a sviluppare organi specializzati per incoraggiare gli animali a disperdere i loro semi. Tutti gli animali sono eterotrofi, devono cioè consumare nutrienti organici per sopravvivere, e in generale quegli individui che hanno un migliore accesso ai nutrienti saranno più sani e più forti e, a loro volta, avranno maggiori possibilità di riprodursi. Questa necessità è stata sfruttata dalle piante, che hanno offerto i loro semi agli animali confezionandoli in un rivestimento nutriente. In questo modo gli animali li ingeriscono e in seguito li depositano lontano nei loro escrementi (zoocoria). Gustose, seducenti e colorate strutture di polpa zuccherina si sono evolute per racchiudere i semi di varie Angiosperme, attirando al banchetto gli animali che, inconsapevoli, li trasportano in luoghi adatti per la loro germinazione. Erano comparsi i frutti, ma questa è un’altra storia che racconteremo un’altra volta.
Grazie a loro, ho scritto:
Benson W (2016) Kingdom of Plants. A Journey Through Their Evolution. Collins.
Gledhill D (2008) The Names of Plants. Cambridge University Press.
Ingrouille M, Eddie B (2006) Plants: Evolution and Diversity. Cambridge University Press.
Thompson JD (2005) Plant Evolution in the Mediterranean. Oxford University Press.



