“Affidati alla forma per vincere sul nemico numeroso. E quest’ultimo non riuscirà a capire come hai fatto. Tutti vedono la forma con cui ho vinto, ma nessuno sa cosa mi porta a decidere la forma della vittoria. Evita di ripetere le tattiche vittoriose del passato, Perché la forma deve essere suggerita dall’infinità varietà delle circostanze.
Il fine del dare forma alle operazioni militari è diventare senza forma. Quando si è senza forma, nemmeno le spie più abili riescono a scoprire nulla e il nemico saggio non avrà elementi per poter preparare i suoi piani.
La forma che vince i molti, non appare ai molti. Dopo la vittoria, la mia forma sarà palese a tutti. Prima della vittoria, nessuno sa la forma che impiegherò.”
da “L’arte della guerra”, Sun Tzu
Era da tempo e in vari posti e occasioni che meditavo sui contenuti di questo post. L’articolo mi ha fatto penare perché le informazioni sul mimetismo nelle piante sono frammentarie, a volte contraddittorie, sembrano di colpo arrestarsi negli anni ’80 per poi ricomparire negli ultimi anni e sono state spesso pubblicate su riviste non specialistiche. Spero almeno di aver fatto un buon lavoro.
Il mimetismo coinvolge un organismo (l’imitatore) che simula segnali provenienti da un altro organismo (il modello), i quali sono percepiti come interessanti da un terzo organismo (l’operatore o meno elegantemente, come spesso si trova in letteratura, “dupe” = “babbeo, pollo”), cosicché l’imitatore ha vantaggi riproduttivi perché l’operatore/pollo lo scambia per il modello. Dopo questo incipit così didascalico, cominciamo con la spiegazione più terra terra. Prima di tutto, il mimetismo non è una prerogativa degli animali ma riguarda anche le piante. Se n’è parlato e scritto di meno soltanto perché il fenomeno è stato spesso associato ai movimenti veloci degli animali o a un fenomeno regolato da un sistema nervoso organizzato, assente nelle piante.
Nei fiori, il mimetismo si esprime ai massimi livelli. Il principio alla base è semplice: imitare le caratteristiche di un fiore modello, il cui habitat si sovrappone a quello dell’imitatore, al fine di trarne vantaggio. La maggior parte dei fiori imitatori non producono affatto nettare ma prendono come modello fiori molto più grati in termini di succulenti carboidrati.
Esempi di mimetismo batesiano (quando una specie sfrutta la sua somiglianza con una specie che vive nel suo stesso territorio), soprattutto nelle orchidee (1/3 di esse, circa 10.000 specie, sono imitatrici), sono comuni e già dopo una rapida ricerca in rete, ne troverete tantissimi che abbracciano la morfologia del fiore, il colore, la riflettanza spettrale, l’architettura delle infiorescenze e il tempo di fioritura. Il vantaggio dei fiori scrocconi consiste nel fatto che hanno più visite da parte dei trasportatori di polline, fornendo però una parva ricompensa in termini di nettare. Di solito, anche i fiori generosi sono parzialmente avvantaggiati dalla maggiore diffusione degli impollinatori ma l’asimmetria di benefici (che caratterizza il mimetismo batesiano) però rimane, in quanto l’imitatore riceve molti più vantaggi. Il mimetismo batesiano può essere intraspecifico (auto-mimetismo, o mimetismo bakeriano, di cui parleremo in seguito) ma molto più spesso è interspecifico, per cui imitatore e modello sono due specie distinte ma simpatriche (vivono nello stesso posto e i loro habitat si sovrappongono) e il modello è di solito più abbondante dell’imitatore. Ad esempio, le orchidee oncidioidi (Oncidiinae; circa 1700 specie tropicali, con vari habitat e dimensioni) presentano fiori di colori e forme che ricordano quelli di altre specie simpatriche, e in particolar modo delle Malpighiaceae, che producono nettare e polline. Ci sono tantissimi altri casi di questo genere nelle orchidee del genere Epidendrum e Ophrys. Questo particolare tipo di mimetismo riproduttivo fiorale può essere anche deinito mimetismo dodsoniano (dal tassonomista, botanico e specialista di orchidee Calaway H. Dodson).
Come viene ingannato il babbeo? Un caso tra tanti: l’orchidea Cephalantera rubra (che non produce nettare) imita varie specie di campanula. A prima vista (foto in basso) la somiglianza non è così evidente ma la prospettiva cambia se a vedere i fiori sono le api. Queste hanno un’estensione della vista fino all’ultravioletto vicino e, come molti insetti, distinguono poco le varie tonalità di rosso-violetto. La forma e il profumo sembrano invece avere meno importanza nella scelta.

L’orchidea Cephalantera rubra (imitatrice; a sinistra) e la campanula Campanula persicifolia (modello; a destra).
Il colore rosso è strategico anche per i fiori impollinati dai colibrì. In questi uccelli, la capacità di discriminare tra varie tonalità di rosso sembra essere molto scarsa (“confusione spettrale”) e nel loro campo visivo il rosso si staglia in maniera netta rispetto allo sfondo verde delle foglie. Per questo motivo, il colore di molti fiori che si servono dell’impollinazione dei colibrì ma che, ingrati, non offrono molte ricompense agli operosi uccellini “converge” verso il rosso. In molte altre specie, il colore rosso è tipico di frutti non ancora maturi, come avviene nella mora, nell’alaterno, nell’artiglio del diavolo, nell’oleandro e in molte specie di pistacchio, i quali in questo stadio possono essere aspri, astringenti, spinosi o velenosi. Si tratta quindi di una colorazione aposematica, che ha il preciso compito di dire ai frugivori: “Faccio schifo, non mangiatemi, almeno fino a quando non sono maturo e potete fungere da vettore per i miei semi”. Per questo, alcuni ipotizzano che altre specie vegetali imitino questo colore, anche non essendo particolarmente disgustosi. In questo modo, risparmiano energia imitando le specie pericolose (mimetismo batesiano anche qui).
Paradigmatico è il caso della passiflora , la quale produce sostanze tossiche nelle sue foglie per respingere gli attacchi di diverse specie di insetti. C’è però una specie di eliconide (Heliconius charithonia) i cui bruchi hanno una saliva in grado di rendere innocui metaboliti tossici prodotti nelle foglie della passiflora. In più, i bruchi usano queste sostanze per produrre essi stessi tossine che li rendono inappetibili ad altri insetti una volta diventati farfalle! Inutile dire che tutte le farfalle di Heliconius preferiscono le foglie di passiflora per deporre i loro grappoli e colonne di uova gialle. E’ a questo punto che la passiflora s’incazza e corre ai ripari producendo sulle foglie delle escrescenze gialle (stipule), molto simili alle uova di eliconide, munite di una piccola ghiandola secernente nettare, di cui formiche e vespe sono ghiotte. Sono queste ultime che, attirate dal dolce, ne approfittano per banchettare a base di nettare, uova (stavolta vere) di eliconide e bruchi. Non saprei come classificare questo tipo di mimetismo, in cui una pianta imita le uova appetitose di un insetto.
Ma il simpatico vegetale non si ferma qui. Il fiore di passiflora è evidentemente molto bello: ha una struttura complessa ed è un caleidoscopio di colori e assomiglia quasi ad una piattaforma di atterraggio a più stadi. A causa dell’aggressività tipica di molte specie di formiche, somigliare ad esse può essere una strategia contro gli erbivori. Questo è un tipo di mimetismo batesiano che ricorda molto quello degli animali, dove una specie innocua trae vantaggio imitando una specie “pericolosa” che presenta una colorazione aposematica (la colorazione di una parte più o meno estesa del corpo di un animale a fini di avvertimento contro possibili predatori). Molte specie di passiflora, infatti, presentano punti e trattini neri che mimano formiche e afidi nella parte interna del fiore (sulle antere), proteggendolo dagli eventuali erbivori (foto in basso).
 Fiore di passiflora con puntini neri sulle antere (raggi al centro del fiore) (Lev-Yadun, 2009).
Fiore di passiflora con puntini neri sulle antere (raggi al centro del fiore) (Lev-Yadun, 2009).
La medesima tattica è utilizzata in altre specie vegetali (nella foto in basso: Xantium trumarium e Alcea setosa) che producono puntini e trattini su fusti e foglie, oppure baccelli immaturi che imitano le colorazioni aposematiche e la forma di bruchi velenosi o irritanti. Nella foto in basso si vedono i baccelli immaturi di una specie di fava (Vicia peregrina) che imitano il bruco irritante tipico dei lepidotteri pieridi.
 Esempi di mimetismo batesiano di formiche, afidi e lombrichi da parte di piante (Lev-Yadun e Inbar, 2002).
Esempi di mimetismo batesiano di formiche, afidi e lombrichi da parte di piante (Lev-Yadun e Inbar, 2002).
Curioso è il comportamento della “pianta semaforo” (o telegrafo) Desmodium motorium (o gyrans), con I suoi misteriosi movimenti fogliari lungo traiettorie ellittiche ad intervalli di pochi minuti. Desmodium sembra mimare il battito della ali delle farfalle o in generali di artropodi alati, scoraggiandoli a deporre le loro uova (“Sono occupata”). Il movimento delle foglie potrebbe anche attrarre una serie di organismi insettivori (uccelli, rettili, mammiferi diurni e artropodi), che in questo modo pattuglierebbero la pianta, ripulendola da eventuali predatori erbivori.
Gli enigmatici movimenti fogliari ellittici ad intervalli regolari di Desmodium motorium, la pianta semaforo.
Sfruttano il mimetismo batesiano al contrario (imitano specie innocue fingendosi inoffensive) alcune piante carnivore come le Nepenthes, che hanno trappole costituite da foglie trasformate a forma di bottiglie o boccette allargate, somiglianti a degli otri, coperte da un labbro superiore. Queste piante tropicali vivono in habitat con acqua, luce e radiazione solare in abbondanza, fotosintetizzano a pieno ritmo e producono molti zuccheri; i loro fiori sono quindi pieni di nettare e attirano quindi molti insetti. Quel che a loro manca è l’azoto, che si procurano facendo scivolare gli insetti sui bordi delle foglie modificate. Risultato: i malcapitati cadono negli otri ripieni di liquido, finendo per essere digeriti da enzimi proteolitici prodotti dalla pianta. Ora, dal momento che i fiori delle Nepenthes producono nettare, che solo una piccola percentuale di insetti finisce nelle trappole, e che sia pianta che insetti hanno i loro benefici, l’inganno è parziale. Sembra essere piuttosto una sorta di mutualismo, periodicamente mortale. Da una visione antropocentrica, possiamo dire che c’è più onestà in questo tipo di interazione.
 Una mosca sull’orlo del Maelström nepenthiano.
Una mosca sull’orlo del Maelström nepenthiano.
Abbiamo accennato prima al mimetismo bakeriano, dal nome del naturalista inglese Herbert G. Baker, una forma di auto-mimetismo (o mimetismo intraspecifico) che avviene in una singola specie. Ad esempio, nelle piante dioecie – che hanno fiori maschili e femminili su piante separate e presentano marcate differenze tra i due tipi di fiori – i fiori femminili (che non producono ovviamente polline) mimano quelli maschili, ingannando così gli impollinatori, altrimenti non sarebbero mai fecondati. Questa strategia è comune in molte specie di Caricaceae, fra cui c’è anche la papaya. Nonostante la differenza morfologia dei fiori dei due sessi, l’impollinazione erronea avviene lo stesso perché le falene che impollinano la papaya (Sphingidae) agiscono all’alba, quando la luce non è diretta ed è più facile confondersi.
Molto frequente tra i fiori è anche il mimetismo mülleriano, in cui non c’è inganno né una chiara distinzione tra modello e imitatore, e tutti gli attori contribuiscono equamente e non a senso unico come nel mimetismo batesiano. Qui le caratteristiche convergenti tra due o più specie che offrono nettare agli impollinatori aumentano la reperibilità della ricompensa per questi ultimi, migliorando così l’impollinazione per tutte quante le specie (“impollinazione cooperativa”). Questo vale soprattutto per i fiori delle specie che offrono meno nettare, i quali si avvantaggiano dell’ “effetto magnete” da parte fiori più generosi (sconfiniamo nel parassitismo?). E’ come trovarsi nel gran ballo delle debuttanti vegetale, in cui tutti i fiori si mettano contemporaneamente il vestito buono per attirare al meglio gli insetti impollinatori di alto rango.
Tornando alle irritanti spine, stavolta delle piante, anch’esse sono un caso di mimetismo mülleriano al contrario dove in questo caso più specie, tutte inappetibili, si imitano a vicenda. Se pensate ai cactus e all’agave dei deserti nord americani, all’aloe, all’euforbia e alle acacie africane, alle tante specie spinose della macchia mediterranea, si può ipotizzare che le loro strutture spinose e le loro colorazioni, per alcuni versi molto simili, siano un preciso segnale per gli erbivori, che in questo devono imparare un unico segnale di avvertimento, anziché uno diverso per ogni specie, diminuendo di conseguenza il numero di individui di ogni specie sacrificati per consentire l’apprendimento del segnale aposematico. Alcuni mettono in dubbio l’esistenza di questo mimetismo perché le caratteristiche simili di molte specie di piante sono sovente dovute al fatto che vivono in habitat simili e sono sottoposte alle stesse condizioni ambientali; quindi sarebbe più giusto chiamarlo evoluzione convergente.
Ci sono però anche casi di animali che imitano le piante! Gli adulti di un coleottero australiano (Procrisina pictipennis) imitano i frutti (capsule) in via di sviluppo della pianta ospite (Leptospermum polygalifolium). La forma del corpo e il colore degli adulti, che si nutrono e si accoppiano in prossimità dei margini dei rami della pianta ospite, fanno sì che i predatori non si accorgano di loro. Addirittura, se disturbati, i coleotteri presenti sulle foglie si lasciano cadere al suolo, proprio come farebbe un frutto maturo!
E ancora, la cavalletta semi-acquatica Paulinia acuminata e la pianta di cui si nutre, la felce fluttuante Salvinia auriculata, entrambe specie tropicali, sembrano aver trovato un compromesso tra mimetismo e convergenza adattativa. Le giovani ninfe della cavalletta (foto in basso a sinistra) hanno lo stesso colore e la stessa struttura superficiale delle foglie della pianta. In questo modo, difficilmente sono disturbate dai predatori. Le ninfe degli stadi successivi e gli adulti (foto in basso a sinistra), più grandi delle foglie ovali di Salvinia, esibiscono un mimetismo somatolitico (cioè, quando i colori e i disegni del corpo sembrano rompere la figura dell’animale, rendendone difficile l’avvistamento) e imitano il confine tra due foglie compresa la loro discontinuità. Infine, sia le giovani ninfe che le foglie sono repellenti all’acqua a causa delle cere epicuticolari, che oltretutto possiedono la stessa ultrastruttura per convergenza adattativa (vivendo in uno stesso ambiente, si finisce a volte per avere alcune caratteristiche simili, come una sorta di analogia funzionale).
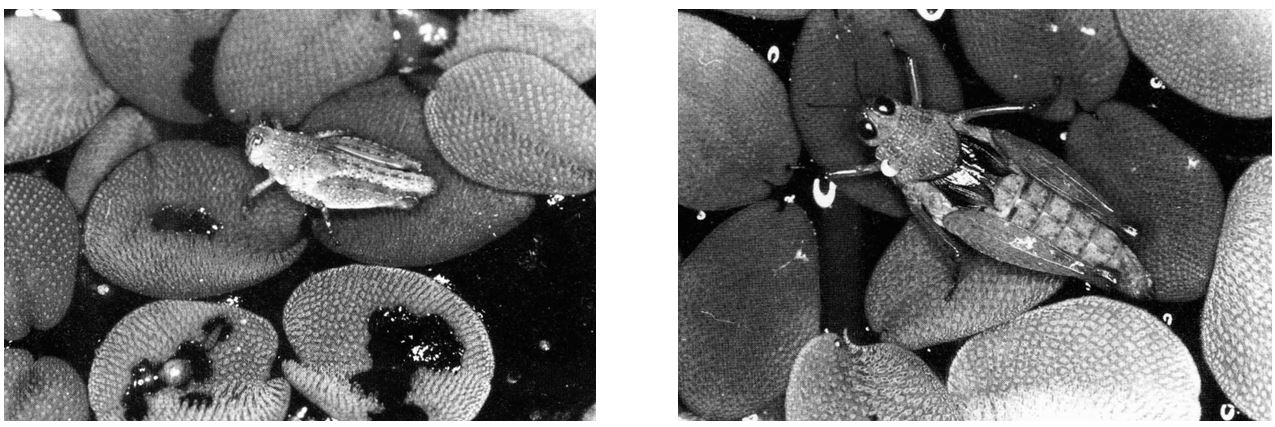 Giovane ninfa (a destra) e adulto (a sinistra) di Paulinia acuminata su foglie di Salvinia auriculata (Barthlott et al., 1994).
Giovane ninfa (a destra) e adulto (a sinistra) di Paulinia acuminata su foglie di Salvinia auriculata (Barthlott et al., 1994).
E poi ci sono fiori che imitano le caratteristiche (odore, colore, forma, consistenza) della carne in putrefazione, come il gigantesco fiore di Amorphophallus titanum (vedete un mio vecchio post qui), in grado di attirare mosche e vermi coprofagi e approfittano di loro per essere impollinati. Molte piante carnivore utilizzano la stessa tattica, basandosi soprattutto sulle sostanze odorose che attirano gli insetti. Un mix di feromoni e forma è un’arma di cui si servono parecchie specie di orchidee del genere Ophrys, i cui fiori, muniti di particolari ghiandole, producono fragranze simili ai feromoni delle femmine degli insetti imenotteri (api, vespe) e ne mimano anche l’addome, attirando irresistibilmente i maschi impollinatori (foto in basso). I maschi sono fortemente attirati dal fiore e cercano di copularlo (pseudocopulazione; video in basso). L’insetto atterra sul labello, il petalo più grande e sporgente e, cercando di accoppiarsi, attiva un meccanismo per cui il fiore cambia conformazione, cosicché le sacche piene di polline che pendono dalla parte superiore del fiore si scontrano con l’addome dell’insetto. Questa strategia, chiamata anche mimetismo pouyanniano, è talmente efficace che si è sviluppata in maniera indipendente nelle orchidee di Australia, Eurasia e Sud America, ed è particolarmente diffusa nei tropici.
 Orchidea specchio (a sinistra), orchidea ape (al centro) e orchidea martello (a destra).
Orchidea specchio (a sinistra), orchidea ape (al centro) e orchidea martello (a destra).
Nonostante la sorprendente ma non perfetta imitazione, gli insetti maschi si lasciano ingannare perché le orchidee fioriscono in sincronia con la loro schiusa, la quale avviene poco prima di quella delle femmine. In un’atmosfera a dominanza prettamente maschile, con un’alta competizione per le poche femmine adulte, i maschi sono sessualmente frustrati e si buttano sulla prima cosa che ricordi anche vagamente un’ape femmina.
Esempi di pseudocopulazione in alcune orchidee.
L’esempio più bello è sicuramente quello dell’orchidea martello (Drakeae spp.), nativa dell’Australia e delle sue vespe impollinatrici (video in basso) ma ci sono anche esempi più nostrani, come quello tra l’orchidea mediterranea Ophrys tenthredinifera e l’ape cornuta Eucera nigrilabris.
L’orchidea martello in azione.
Fuori da tutti i tipi di mimetismo sopra elencati sono le risposte dirette delle piante ad un pericolo diretto costituito da un predatore. Alcune specie di piante ricordano la forma, le dimensioni, il colore e persino il disegno di oggetti inanimati, come avviene nelle “piante sasso” del genere Lithops, originarie del Sud Africa, le quali sono impossibili da distinguere da sassi e ciottoli, soprattutto durante la stagione secca, quando si disidratano e vengono in parte coperte dalla sabbia. Persino la loro fioritura è brevissima e avviene solo durante la stagione umida, quando l’erba è abbondante per gli erbivori e il pericolo di essere mangiate diminuisce. In questo caso, l’effetto della pressione selettiva di uccelli, piccoli mammiferi e ungulati erbivori, che una volta solcavano in abbondanza le pianure africane, ha compiuto il miracolo di trasformare una pianta in un sasso.
Le piante sasso “in azione”.
Mimetismo definitivo, senza troppe definizioni, è quello di altre due piante. La prima si finge morta come un soldato spacciato. Si tratta della Mimosa pudica, di cui ho già scritto in un altro post, la quale richiude le pinnule che compongono le sue foglie in seguito ad un lieve tocco. Se il contatto è più sostenuto, sia le foglie che i rametti della mimosa si accasciano, e volte l’intera pianta si stende al suolo, sparendo alla vista del predatore. La seconda è la Monotropsis odorata, incapace di compiere fotosintesi ma in grado di ricavare sostentamento da un fungo con cui vive in simbiosi. La pianta è caratterizzata da fusti e boccioli colorati di rosso vivo e circondati da brattee. Queste ultime, una volta seccate, assumono una colorazione marroncina simile a quella della lettiera di foglie, cambiando la livrea della pianta, che passa inosservata e confonde i predatori. Inutile dire che rimuovere le brattee della pianta significa condannarla ad una maggiore pressione predatoria.
 Monotropsis odorata con le brattee intatte (a sinistra) e con le brattee rimosse sperimentalmente (a destra) (Klooster et al., 2009).
Monotropsis odorata con le brattee intatte (a sinistra) e con le brattee rimosse sperimentalmente (a destra) (Klooster et al., 2009).
Al contrario delle piante sasso, che sfruttano una pressione selettiva del passato, ci sono altre piante che invece sfruttano la pressione selettiva attuale attuata dall’Homo sapiens agricolo. Sono bastati poco più di 10.000 anni perché alcune erbe infestanti abbiano “imparato” ad assumere una o più caratteristiche di una pianta domesticata mediante generazioni di selezione artificiale. Questo mimetismo – giuro che è l’ultima definizione della giornata – è chiamato vaviloviano (oppure anche “mimetismo della coltura” o “mimetismo della malerba”. Il nome proviene dal genetista Nikolai Vavilov, il quale negli anni ’20 osservò che nei campi di frumento, le erbacce che somigliavano alla segale e all’avena venivano raccolte e selezionate accidentalmente dall’uomo per alcuni tratti tipici della domesticazione, quali i semi grandi, le pannocchie rigide e il portamento eretto, diventando talvolta colture utili e interamente dipendenti dall’uomo per la loro sopravvivenza. Certo, per l’agricoltore il problema è insidioso, dal momento che deve distinguere il malefico mimo infestante ad occhio e rimuoverlo a mano. L’avvento dell’agricoltura è stata una manna dal cielo per le erbe imitatrici, soprattutto perché il massimo grado di somiglianza si verifica prima della fioritura, quando la competizione con la specie coltivata è più alta (una volta fiorita diventa di solito più identificabile ma il danno è stato ormai fatto). I modelli più ambiti sono ovviamente quelli più diffusi: i cereali. Nel riso, ad esempio, è difficilissimo distinguere le specie selvatiche improduttive che crescono insieme a quella coltivata, anche perché il periodo di fioritura e di germinazione sono molto simili e l’ibridizzazione tra le varie specie è molto frequente. Sempre nel riso, altri due mimi sono Echinochloa oryzoides (foto in basso) e Echinochloa phyllopogon, due malerbe obbligate originarie dell’Asia ma oggi divenute un problema internazionale risolvibile in parte e temporaneamente solo con l’adozione di varietà di riso precoci, il cui seme matura prima di quello dell’infestante. Anche in Italia, l’areale di distribuzione di queste due specie infestanti è limitata al nord-ovest (vedete qui), dove difatti viene coltivato il riso.
 Il mimo Echinochloa oryzoides (a sinistra) e il suo modello, il riso coltivato (Oryza sativa) (a destra).
Il mimo Echinochloa oryzoides (a sinistra) e il suo modello, il riso coltivato (Oryza sativa) (a destra).
La pressione selettiva dell’uomo ha promosso anche un altro tipo di mimetismo, quello dei semi. I semi di veccia comune (Vicia sativa), leguminosa parente della fava, si sono probabilmente trasferiti con quelli della lenticchia coltivata (Lens culinaris) dalla Mezzaluna Fertile al Medio Oriente e poi in tutto il Mediterraneo. Per somigliare sempre di più alla lenticchia, i semi di veccia hanno ridotto il periodo di dormienza, aumentato la loro abilità competitiva e la biomassa e si sono adattati a nuovi ambienti, finendo per essere domesticati come la lenticchia e raccolti insieme ad essa. Il misfatto non sembra essere un caso isolato perché nelle giare piene di sementi risalenti anche a 5000-6000 anni fa sono state trovate miscele di cicerchia (Lathyrus sativus), lenticchia (Lens culinaris) e pisello (Pisum sativum), dove stavolta l’intrusa era la cicerchia, in seguito domesticata. Una situazione molto simile è stata riscontrata nel mimetismo dei semi di Camelina sativa (varietà linicola) con quelli del lino. Camelina cresce nei campi di lino e, sottile, con un fusto non ramificato, pallida, somiglia ad esso. I suoi semi maturano contemporaneamente a quelli del lino e, poiché i frutti rimangono attaccati alla pianta, sono raccolti insieme a quelli della coltura principale. Di solito, i semi diversi dal lino sono rimossi durante i processi di trebbiatura e vagliatura, ma quelli di Camelina hanno proprietà fisiche simili a quelli del lino (ad occhio sembrano invece; foto in basso) e sono lanciati alla stessa distanza dalle macchine per la vagliatura. Così, i semi delle due specie finiscono per essere ripiantati insieme l’anno successivo; e l’inganno si perpetua.
 Il seme mimo della Camelina sativa (a sinistra) e il suo modello, il seme di lino (Linum usitatissimum) (a destra).
Il seme mimo della Camelina sativa (a sinistra) e il suo modello, il seme di lino (Linum usitatissimum) (a destra).
Grazie a loro, ho scritto:
Ackerman JD (1986) Mechanisms and evolution of food-deceptive pollination systems in orchids. Lindleyana 1 (2): 108-113
Barrett SCH (1987) Mimicry in plants. Scientific American 255 (9): 76-83
Barthlott W, Riede K, Wolter M (1994) Mimicry and ultrastructural analogy between the semi-aquatic grasshopper Paulinia acuminata (Orthoptera: Pauliniidae) and its foodplant, the water-fern Salvinia auriculata (Filicatae: Salviniaceae). Amazoniana 8 (1/2): 47-58
Benson W (2010) Kingdom of Plants. A Journey Through Their Evolution. Kew Royal Botanic Gardens.
Bleiweiss R (1990) Spectral confusion by hummingbirds and the evolution of red coloration in their flowers: a new hypothesis (Müllerian mimicry/wavelength discrimination/shade/color name). Transactions of the Wisconsin Academy of Sciences, Arts and Letters 78: 33-38
Dupre D (2011) Evidence for floral mimicry in Epidendrum radicans (Orchidaceae) with Asclepias curassavica (Apocynaceae) and Lantana camara (Verbenaceae). PhD thesis, University of Florida. http://digital.lib.usf.edu/SFS0001543/00001
Erskine W, Smartt J, Muehlbauer FJ (1994) Mimicry of lentil and the domestication of common vetch and grass pea. Economic Botany 48 (3): 326-332
Harder LD, Barrett SCH (2006) Ecology and Evolution of Flowers. Oxford University Press.
Hawkeswood TJ (2002) Mimicry of host plant fruits by adults of the Australian leaf beetle Procrisina pictipennis (Boheman) (Coleoptera: Chrysomelidae). Mauritiana (Altenburg) 18 (2): 239-243
http://en.wikipedia.org/wiki/Mimicry_in_plants. Mimicry in plants
http://gianlucagrossi.blogspot.it/2010/01/piante-mimetiche.html. Piante mimetiche
Joel DM (1988) Mimicry and mutualism in carnivorous pitcher plants (Sarraceniaceae, Nepenthaceae, Cephalotaceae, Bromeliaceae). Biological Journal of the Linnean Society 35 (2): 185-197
Klooster MR, Clark DL, Culley TM (2009) Cryptic bracts facilitate herbivore avoidance in the mycoheterotrophic plant Monotropsis odorata (Ericaceae). American Journal of Botany 96 (12): 2197-2205
Lev-Yadun S (2013) The enigmatic fast leaflet rotation in Desmodium motorium: Butterfly mimicry for defense? Plant Signaling and Behavior 8 (6): e24473
Lev-Yadun S, Ne’eman G, Izhaki I (2009) Unripe red fruits may be aposematic. Plant Signaling & Behavior 4 (9): 836-841
Lev-Yadun S (2009) Ant mimicry by Passiflora flowers? Israel Journal of Entomology 39: 159-163
Lev-Yadun S (2009) Müllerian mimicry in aposematic spiny plants. Plant Signaling & Behavior 4 (6): 482-483
Lev-Yadun S, Inbar M (2002) Defensive insect mimicry in plants. Biological Journal of the Linnean Society 77: 393-398
Powell MP, Pupulin F Warner J, Chase MW, Savolainen V (2003) Flora mimicry in oncidioid orchids. Lankesteriana 7: 109-110
Schildknecht H (1986) A Chemical Power of Movement in Higher Plants. Natural Product Chemistry: 358-382