Superbo muschio

Le Briofite (muschi) viventi sono rappresentate da circa 900 generi e quasi 24.000 specie. Non sono una parte cospicua della flora della Terra ma, nelle foreste pluviali, la biomassa vivente di muschi e epatiche può superare quella di tutte le altre piante. I muschi del genere Sphagnum, estremamente leggeri e permeabili, coprono circa l’1% della superficie terrestre e sono tra le piante più grandi della terra.
I muschi richiedono elevate quantità di acqua per compiere il loro ciclo vitale, e la maggior parte delle specie copre superfici non porose o semiporose, come rocce, alberi e strutture artificiali. In generale, i muschi abbondano in aree umide e ombreggiate, ma possono anche sopravvivere a lunghi periodi di disidratazione, ad esempio in deserti e tundre, e ne sono stati trovati anche nelle zone costiere dell’Antartide. Generalmente, hanno una capacità molto limitata di controllare l’assorbimento e la perdita d’acqua e la maggior parte delle specie è costituita da piccole piante, molte delle quali di altezza minore a 2 cm. Tra le prime piante pioniere a colonizzare la terraferma (a partire da 470 milioni di anni fa) ci furono proprio gli antenati degli attuali muschi, che per crescere estraevano, come fanno ancora oggi, minerali come calcio, magnesio, fosforo e ferro dalle rocce, alterando in tal modo anche il ciclo globale del carbonio, e quindi il clima, e aprendo la strada a specie colonizzatrici successive. Mi ha fatto sempre specie pensare a questa primordiale distesa verde avanzante su una terraferma che era praticamente priva di suolo e di vita, rocciosa e scabra.
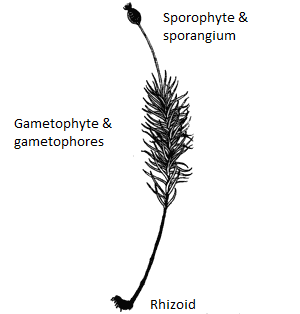
Gametofito e sporofito di Dawsonia superba.
I muschi differiscono dalle piante vascolari vere dall’assenza di tessuto vascolare e dalla presenza di un gametofito (l’individuo che darà origine agli spermi, se maschio, o all’uovo, se femmina) indipendente dal punto di vista nutrizionale nel loro ciclo vitale. Il gametofito è a sessi separati, aploide (con metà del corredo genico). Quando gli spermi, veicolati dall’acqua, fecondano l’uovo del gametofito femminile, si origina lo sporofito diploide. Quest’ultimo crescerà sullo stesso gametofito femminile e darà origine alle spore. È facile riconoscere lo sporofito dal gametofito femminile perché si ha una forma a filamento allungati che termina con una capsula arrotondata e si erge dal “tappeto” verde sottostante di gametofiti. Rispetto alle piante superiori, dove lo sporofito domina sul gametofito, la situazione è quindi rovesciata.
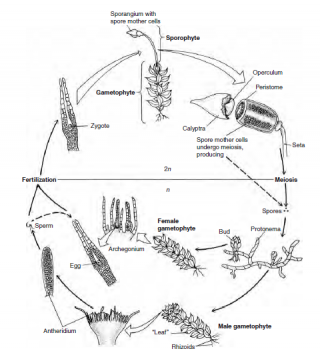
Ciclo vitale di un muschio.
I muschi non sono piante vascolari (non hanno xilema e floema), e le loro foglie (filloidi) non sono vere foglie ma sono comunque specializzate per il trasporto e la conservazione dell’acqua. Un’altra caratteristica che li accomuna è l’assenza di stomi, presenti solo in alcuni dei loro sporofiti.
Le foreste pluviali sono particolarmente ricche di muschi, epatiche e licheni: il muschio Dawsonia superba (fino a 50 cm di altezza, anche se alcuni dicono di averne trovate fino a 70 cm), l’epatica fogliosa Plagiochila stephensonii (alta 25 cm), l’epatica frondosa Monoclea forsteri (alta 20 cm) e il lichene epifita Pseudocyphellaria coronata (larga 20 cm) sono esponenti incredibilmente grandi per ogni gruppo e sono molto diffusi in Nuova Zelanda, ma anche in Australia e Nuova Guinea.

Tappeti di Dawsonia superba.
In particolare, il muschio Dawsonia superba, il più grande muschio vivente, a uno sguardo disattento somiglia ad una piantina di pino, con la quale spesso viene confusa. Il fusto principale non è ramificato e porta foglie numerose e sottili di circa 20-35 mm di lunghezza, ognuna delle quali ha una guaina oblunga alla base che si restringe immediatamente in una lamina lunga e dentata (con 40-60 lamelle). Le lamelle sono alte 4-6 cellule, con le cellule terminali più larghe e ispessite di quelle sottostanti. Questo muschio (insieme ad altri muschi dei generi Polytrichadelphus, Atrichum, Polytrichum e Dendroligotrichum) cattura e trattiene l’acqua piovana sulla superficie superiore delle foglie. Si potrebbe pensare che l’acqua scorra giù ma invece è ancorata per capillarità sulla nervatura centrale della foglia (anche se parlare di “foglia” e “nervatura” per un muschio è alquanto improprio, ma giusto per rendere l’idea). Sulle nervatura centrale, infatti, ci sono file parallele di sottilissime lamelle. Una piccola porzione di acqua bagna gli spazi capillari tra le lamelle, e poi quest’acqua trattiene il resto dell’acqua per coesione (l’attrazione delle molecole di acqua tra di loro). In alcune specie di Dawsonia, la foglia direziona e intrappola l’acqua verso una specie di cilindro impermeabile, dove l’acqua non viene più persa per evaporazione. Inoltre, un rivestimento di cutina, una sostanza impermeabile, evita le perdite di acqua dalla pagina inferiore della foglia e dalla parte terminale delle lamelle. Nella sezione trasversale di Dawsonia superba, qui in basso, le foglie interne blu sono immature, mentre quelle rosse esterne sono mature. Su tutte, si possono vedere le lamelle parallele strettamente impilate lungo la nervatura centrale della pagina superiore della foglia. Le lamelle, oltre a catturare e trattenere acqua, sono anche fotosintetiche, e quindi aumentano considerevolmente la superficie dei muschi per intrappolare la luce.
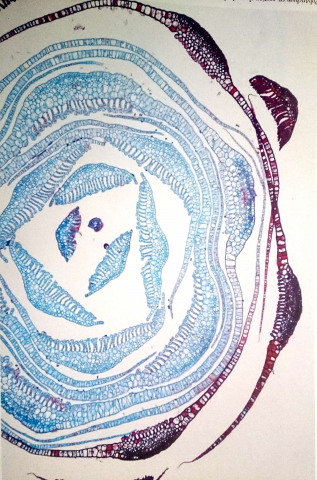
Sezione trasversale di Dawsonia superba.
Una capsula dello sporofito di un muschio contiene almeno migliaia di spore, e alcune letteralmente milioni (65 milioni in Dawsonia, che ha spore piccole e capsule grandi). Fino a quando le spore non sono completamente mature, sono trattenute nella capsula da un solido coperchio chiamato opercolo. Appena sotto l’opercolo c’è un anello (a volte due) di dentini chiamato peristoma. Dopo che il coperchio cade, il peristoma controlla la dispersione delle spore: i dentini, una volta bagnati, si arricciano velocemente nella capsula, per poi raddrizzarsi altrettanto velocemente quando sono asciutti. In questo modo, durante i veloci cambi di umidità dovuti ai venti che spirano nel sottobosco, le spore nella capsula sono lanciate e rilasciate in aria. In Dawsonia superba (insieme ai piccoli muschi del genere Totula), i dentini sono sostituiti da lunghi filamenti arricciati e le spore sono lanciate fuori in aria mediante questi filamenti quando la capsula viene fatta vibrare dal vento o alla sollecitazione fisica dovuta al passaggio di insetti e animali. La capsula larga e piatta di Dawsonia ha la particolarità di avere un denso ciuffo di peli (detti peli peristomiali) lunghi e sottili all’apertura, con opercolo e caliptra (una specie di cappuccio che ricopre la capsula e che deriva dal gametofito originario) entrambi conici. La caliptra è inoltre coperta da peli, da cui il nome “Polytrichaceae”, la subclasse a cui il genere Dawsonia appartiene.

Peli peristomiali e spore (in verde) di Dawsonia superba (150X).
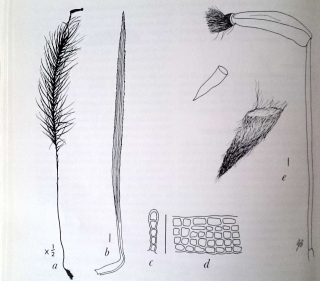
Dawsonia superba: a. pianta intera; b. filloide; c. sezione trasversale della lamella; d. sezione longitudinale della lamella; e. capsula, opercolo e caliptra (secca). Ingrandimento: b. e. 1 mm; c. d. 50 µm.
Al contrario di molti altri muschi, le Polytrichaceae hanno un sistema conduttore interno abbastanza specializzato. Gruppi specializzati di cellule – gli idroidi che trasportano acqua, e i leptoidi che trasportano i prodotti della fotosintesi – sono situate al centro del fusto, e in alcuni casi connettono questo con le nervature delle foglie. Questi tessuti sono strutturalmente simili allo xilema e al floema delle piante superiori. I tessuti conduttori, seppure primitivi, permettono a questi muschi di essere abbastanza alti e dal fusto relativamente spesso perché i prodotti della fotosintesi possono essere traportati facilmente al resto della pianta, e l’acqua e i nutrienti non sono assorbiti solo dalla superficie delle foglie ma anche dalle radici, per poi essere trasportati verso l’alto dai vasi conduttori.
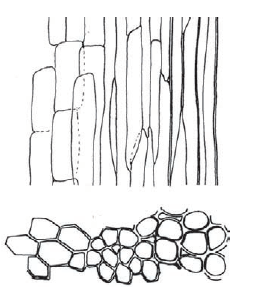
Sezione longitudinale e trasversale degli elementi conduttori in Dawsonia spp.
Al posto delle radici, I muschi come Dawsonia hanno una rete estesa di rizoidi capillariformi, coperti da piccole papille, i quali si sviluppano dall’epidermide e sono in grado di assorbire e traportare l’acqua al resto della pianta.
È impossibile non riconoscere Dawsonia superba per via delle sue dimensioni, ma trovarla per la prima volta in habitat naturali è entusiasmante per ogni botanico o fisiologo vegetale che sia appassionato. Personalmente, sono rimato sbalordito dai tappeti di Dawsonia superba che si stendevano sotto i miei piedi, nel sottobosco della foresta pluviale: di primo acchito, non avrei mai pensato a un muschio ma poi mi sono convinto anch’io!
Grazie a loro, ho scritto:
Beever J (1992) The Mosses of New Zealand. K.W. Allison & John Child, Dunedin, New Zealand.
Ingrouille M, Eddie B (2006) Plants: Evolution and Diversity. Cambridge University Press, Cambridge, UK.
Malcolm B, Malcolm N (1989) The Forest Carpet. Ed. Craig Potton, Nelson, New Zealand.
Taylor TN, Taylor EL, Krings M (2009) Paleobothany. The Biology and Evolution of Fossil Plants. Academic Press, London, UK.
Wardle P (1991) Vegetation of New Zealand. Cambridge University Press, UK.
http://www.lescienze.it/news/2012/02/01/news/piante_clima_glaciazioni_estinzioni_di_massa_fiumi_alveo-827009/












