Il frassino

Mi rendo conto che in questo periodo sto trascurando un po’ i post naturalistici e quindi sono andato alla ricerca di veccie foto, imbatendomi in questa. In una bella escursione di qualche anno fa sul monte Sparviere, tra Basilicata e Calabria, ci imbattemmo due volte in un albero (foto sopra) che, a prima vista ci era sembrato un sorbo. In realtà si trattava addirittura di un gioiello dei nostri boschi: il frassino maggiore (Fraxinus excelsior L.).
Non mi addentro subito in dettagli botanici e fisiologici, dal momento che nei paesi nordici, prima dell’avvento del solito Cristianesimo – quando gli alberi erano probabilmente più rispettati rispetto ad oggi – il Frassino maggiore veniva adorato come pianta sacra: Odino, il più grande degli dei nordici, creò addirittura il primo uomo sulla terra con il legno di frassino . Era poi convinzione comune che bruciare legna di Frassino allontanasse gli spiriti maligni.
In realtà alcuni spiriti maligni li caccia davvero perché, con le foglie raccolte quando sono appena nate, ancora appiccicose e zuccherine, e successivamente essiccate, si prepara un ottimo tè. In fitoterapia, inoltre, si usano semi foglie, linfa, corteccia dei rami giovani perché contengono glucosidi, zuccheri, resine, acido malico, vitamine C e del gruppo B, e hanno proprietà astringente, diuretiche, lassative, sudorifere, toniche.
In Italia è diffuso al settentrione, è presente nei boschi freschi di forra anche nell’Italia centrale ed è raro in quella meridionale. In Basilicata, il Frassino maggiore è segnalato in poche località (alcune zone del Pollino e Monte Pierfaone) ma, almeno io, non l’ho mai visto là. Sul monte Sparviere, invece, l’ho visto per ben due volte. La prima volta in veste di vera e propria apparizione, dal momento che era ai bordi di un precipizio, immerso nella nebbia autunnale, verdissimo e rigoglioso.
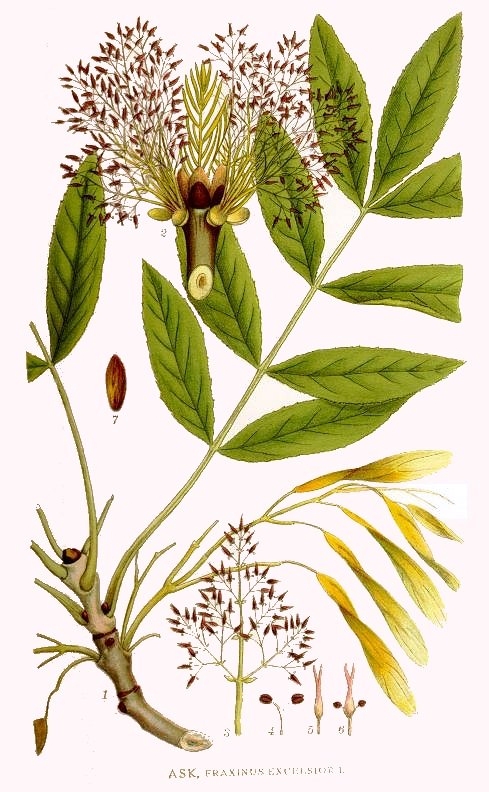
Davvero bello per il portamento e la chioma rada ed elegante. E’ una delle latifoglie più nobili dei nostri boschi. Secondo un detto scandinavo: se la Quercia è il re, il Frassino è l’imperatore. Può essere monoecio (con fiori maschili e femminili separati, oppure con fiori ermafroditi) o monoecio (con fiori maschili o femminili su piante diverse ). Può raggiungere i 40 m di altezza e 1 m di diametro, presenta un rapido accrescimento ed è abbastanza longevo.
Ha un tronco dritto e cilindrico. La corteccia, dapprima liscia e olivastra, successivamente diventa grigio-brunastra e screpolata longitudinalmente. Ha grandi foglie caduche composte imparipennate formate da 9-11 paia di foglioline sessili opposte e minutamente seghettate di colore verde cupo e lucente sulla pagina superiore, più chiare su quella inferiore.
I fiori sono riuniti in pannocchie dense. L’impollinazione è anemofila (mediante il vento), il frutto è una samara monosperma (con un solo seme), lunga 3-4 cm a maturazione, bruno chiaro, obovato- lineare, più o meno arrotondata alla base. Matura a settembre-ottobre e porta un seme che a lungo dormiente a causa di un complesso sistema di inibizione della germinazione.
Il suo legno ad alburno bianco-roseo e durame bruno chiaro con riflessi madreperlacei, è semiduro e di facile lavorazione. Raccoglie in sè molte caratteristiche: morbido eppure solido, mutevole eppur resistente, grazioso eppur forte, lineare, ma rotondo. Si tratta di un legno pregiato, duro, tenace ed elastico, molto ricercato per falegnameria ed ebanisteria (mobili, attrezzi sportivi, come gli sci, mazze da golf, stecche da biliardo, manici di scuri, pioli, zappe, ecc.). Con i polloni si fanno i cerchi da botte e pali per la vite. Da esso si ricava il cosiddetto “ebano grigio” molto ricercato per ebanisteria fine. Viene molto ricercato anche per la fabbricazione di remi, alberi di imbarcazioni e manici di utensili, ed è anche (sprecato!) un buon combustibile.
E’ un albero molto esigente e predilige suoli profondi non compatti, freschi ma anche umidi, da sub-acidi ad alcalini. Tende ad associarsi con l’acero montano, carpino e querce a basse quote, mentre più in alto si associa sia al faggio che all’abete bianco e rosso. E’ un buon bevitore e quindi l’acqua è il suo fattore limitante in quanto ne consuma molta per la sua velocità di crescita e per l’elevata evapotraspirazione: anche piccoli periodi di intensa siccità possono provocare la sua morte; invece sopporta anche per mesi periodi di sommersione delle radici.
E con questo, vado a dormire.
Grazie a loro, ho scritto:
“Latifoglie nobili dei nosti boschi” quaderni di Monti e boschi di G. Bernetti e M. Padula, Edagricole 1984
Fiori e piante del Parco del Pollino. Liliana Bernardo (Ed. Prometeo).
Tree Guide. Owen Johnson & David More. Collins.
“Elementi di fitosociologia” di A. Pirola, CLUEB 1999
http://www.ecomuseovajont.it/laboratorio-telematico/alberi/fraxinus-excelsior-l-frassino-maggiore
http://it.wikipedia.org/wiki/Fraxinus_excelsior
Appunti personali