Piantate a terra

Sulla definizione scritta qui in alto ci sarebbe molto da discutere. Mi soffermerei qui su “makes its own food by photosynthesis” (“produce il suo alimento per mezzo della fotosintesi”), il che è indubbiamene vero, ma solo in parte. Cos’è infatti il cibo per una pianta? I carboidrati che produce per fotosintesi, i minerali del terreno, l’acqua, la luce o altro? Le piante sono organismi fotoautotrofi, per cui la loro fonte di carbonio è inorganica (CO2) (autotrofi) ma la loro fonte di energia è la luce (fototrofi). In un bel libro di Claudio Longo (“Didattica della Biologia”, Ed. La Nuova Italia) ricordo che l’acqua era descritta come alimento per la pianta (“Le piante mangiano acqua, oltre che berla”), e anche questo è vero perché la maggior parte dell’acqua assorbita dalle radici dà turgore alla pianta ma una quantità che si aggira intorno all’1% viene “usata” come fonte di elettroni e protoni per fare fotosintesi (e l’ossigeno di scarto finisce in atmosfera).
Fino a quanto la vita vegetale è stata confinata in acqua, i problemi di approvvigionamento erano limitati: di acqua ce n’era in abbondanza, la CO2 era disciolta in acqua sebbene a concentrazioni minori di quella atmosferica (ma il contrario avveniva per l’ossigeno), in ambienti palustri e costieri c’era non dico abbondanza ma perlomeno non carenza di nutrienti. Nel momento in cui le piante sono uscite allo scoperto (circa 500 milioni di anni fa), sono cominciati i problemi. Una bella definizione di questi problemi l’ho trovata in “Interazioni Pianta-Ambiente” (Sanità di Toppi L. ed altri, Ed. Piccin): “Dipendendo infatti da risorse molto disperse, quali ioni nutritivi, CO2 e luce, le piante sono obbligate a mantenere un’estesa superficie di contatto con l’ambiente esterno”. Per questo motivo le piante sono immerse “testa e piedi” nell’ambiente in cui vivono, la loro chioma ramificata è immersa nell’aria, le loro radici, anch’esse a loro modo “ramificate”, nel terreno.
Questa loro immersione totale nell’ambiente vitale è principalmente dovuta al fatto che la pianta, al contrario di un animale, non può cambiare posto – è un organismo sessile – per cui deve per forza di cose sfruttare al meglio quello che trova intorno a sé. Ciò dà numerosi vantaggi ma è anche un punto debole: a contatto con l’aria, le piante tendono a disidratarsi; e l’acqua mantiene la pianta turgida ma è anche il mezzo indispensabile affinché le cellule spermatiche raggiungano la cellula uovo. Questa “super esposizione” fa sì anche che le piante siano più soggette a temperature estreme, eccessi di luce, competizione con altre piante della stessa o di altre specie, danni causati da parassiti, erbivori e patogeni. Nonostante tutto, tornando al testo di prima, il bilancio è positivo: “La convenienza della scelta compiuta è dimostrata dal fatto che la produzione complessiva di sostanza organica da parte degli ecosistemi terrestri è superiore a quella degli oceani, benché le terre emerse coprano solo un terzo della superficie totale della Terra e l’ambiente acquatico sia in generale molto più favorevole alla vita rispetto all’ambiente subaereo”.
Gli animali terrestri, al contrario, a causa dei loro movimenti, non hanno necessità di radici e foglie. Anche se nell’arte abbiamo bellissimi esempi su questo tema (vedi qui sotto i particolari di piedi-radici e braccia-chioma di Dafne; Apollo e Dafne, Gian Lorenzo Bernini, 1622-1625), gli animali hanno per lo più superfici assorbenti interne (es. mucosa intestinale, alveoli polmonari). C’è qualche eccezione, come nel caso della respirazione cutanea delle rane ma, in linea di massima, gli animali terrestri non scambiano materia attraverso superfici esterne.

Le piante devono quindi interagire completamente con l’ambiente e, per fare ciò, la forma dei loro organi e delle loro cellule assorbenti deve essere ramificata, ricca di introflessioni e di estroflessioni, affinché aumenti l’area di scambio senza che ciò comporti una variazione del volume (vedi questo vecchio articolo). Il meccanismo è sempre il medesimo ad ogni scala: ogni superficie che ha funzione assorbente tende ad aumentare il rapporto superficie/volume mantenendo un volume piò o meno costante e al contempo ramificandosi: “deve esservi una superficie esterna che consenta uno scambio ottimale con l’ambiente al di fuori, ma allo stesso tempo non devono essere raggiunti volumi eccessivi, non potendo la cellula mantenere un fabbisogno metabolico sproporzionato alle sue effettive capacità di scambio” (Il rapporto superficie/volume nelle interazioni del vivente; Francesca Capparelli).
Quando una cellula si accresce, il suo volume aumenta molto più rapidamente della superficie (per esempio, raddoppiando il raggio di una sfera, il volume aumenta di otto volte, mentre la superficie aumenta di quattro volte). Perciò, in una cellula di notevoli dimensioni, la superficie attraverso la quale avvengono gli scambi con l’ambiente circostante non sarebbe in grado di garantire gli scambi nutritivi adeguati alla nuova massa cellulare. Per questo motivo, le cellule si dividono: proprio perché due cellule piccole hanno più superficie di una grande dello stesso volume, e di conseguenza il rapporto S/V aumenta.
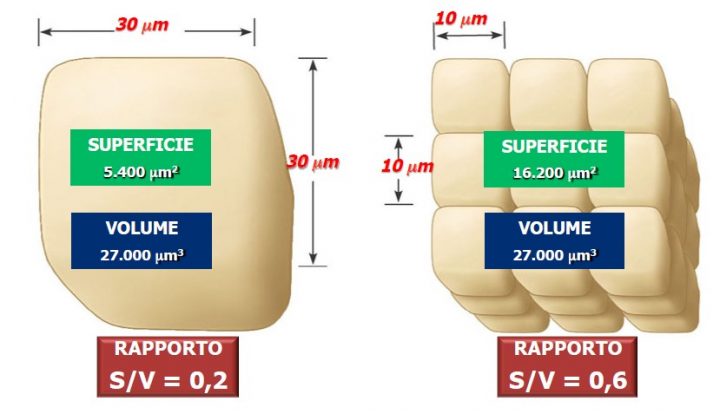
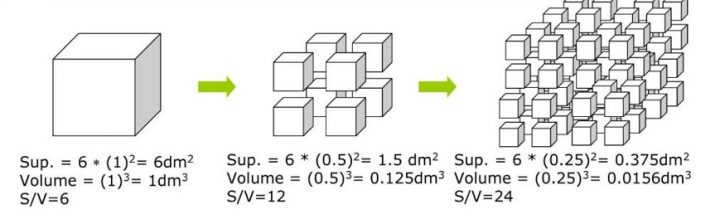
Se approssimiamo la cellula ad un cubo (con la sfera i calcoli sono un po’ più complessi), ci rendiamo subito conto che S/V aumenta quando spezzettiamo il cubo in cubetti più piccoli.
È per questo motivo che le argille hanno grandi superfici se confrontate con pari volumi di sabbia o di limo. Le particelle di argilla, a causa delle loro piccole dimensioni (< 2 µ) hanno un rapporto S/V altissimo e quindi una superficie assorbente enorme, ad es. argille come la montmorillonite e la smectite possono raggiungere 600-800 m2 per g di peso: praticamente la superficie di un campo di calcio in un solo grammo di argilla! Le particelle argillose, proprio per le loro piccole dimensioni e per le cariche negative superficiali, trattengono grandi quantità di ioni di carica positiva utili alle piante, quali calcio, magnesio, potassio e ammonio.
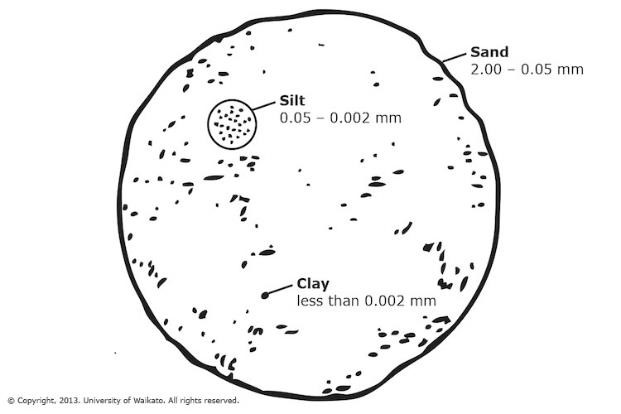
Confronto tra dimensioni di sabbia (sand), limo (silt) e argilla (clay), le principali componenti inorganiche del suolo (Copyright 2013, University of Waikato, Nuova Zelanda).
La stessa cosa avviene se, invece di dividere un solido, lo sfilacciamo e lo ramifichiamo. Anche noi rientriamo in questo caso: pensate alla mucosa intestinale (200 m2 in media di superficie assorbente per “soli” 7 m di lunghezza) grazie a estroflessioni della mucosa chiamate villi e a estroflessioni cellulari, i microvilli. Un altro esempio sono le ramificazioni del sistema respiratorio fino ad arrivare ai bronchi, ai bronchioli e agli alveoli polmonari, strutture concamerate di forma sferica. Grazie alle ramificazioni, i polmoni hanno un peso medio di soli 620-680 g e contengono mediamente 3500-3700 cm3 di aria, ma hanno una superficie alveolare di circa 130 m2. Fenomeni molto simili avvengono nelle strutture a foce dei capillari sanguigni, dove torrente ematico arterioso e venoso si incontrano e avvengono gli scambi di ossigeno, nutrienti e sostanze di scarto con le cellule.
Ogniqualvolta è necessario uno scambio di materia, la superficie aumenta a spese del volume. Negli animali il rapporto S/V è importante anche a livello di organismo: ad es., dall’equatore verso i poli, la dimensione media degli animali aumenta per questioni di ottimizzazione della termoregolazione (legge di Bergmann). Gli animali più grandi, infatti, hanno un rapporto S/V inferiore rispetto agli animali di piccola taglia e quindi disperdono il calore con molta lentezza, trovandosi favoriti in zone con climi freddi. Inoltre, in climi freddi gli animali tendono ad assumere una forma più globosa, dal momento che per questo solido la superficie cresce meno all’aumentare del volume (S/V = 3/raggio) rispetto ad altri solidi (vedi figura in basso).
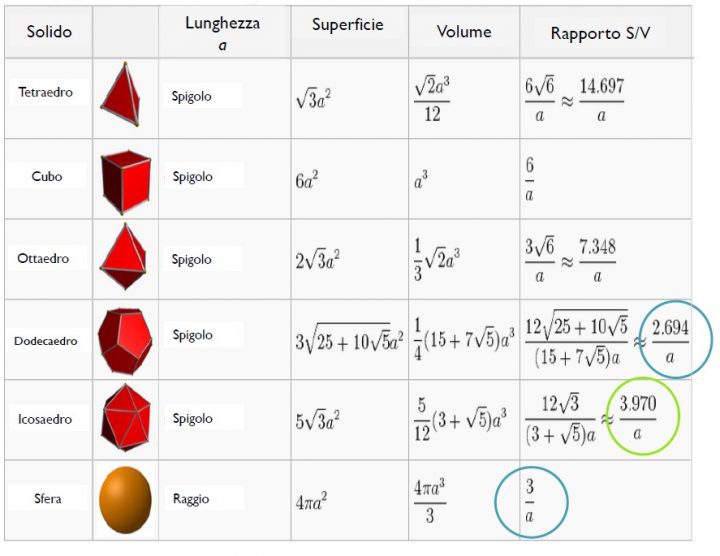
Da: “Rapporto Superficie/Volume”, Fabio Gigante ed Emanuela Montepeloso.
E le piante? La loro superficie assorbente per eccellenza sono le radici per l’acqua e i nutrienti, e le foglie per la luce e la CO2. Anche se ci sono diversi tipi di radici, tutte sono contraddistinte dall’essere ramificate. Una singola pianta di frumento ha una superficie radicale di circa 700 m2! Molte radici sono poi colonizzate da funghi a formare simbiosi (micorrize), e i filamenti di questi funghi (ife) non fanno che aumentare ancora di più la superficie assorbente delle radici, anche fino a 10 volte.

(a sinistra) radice di piantina di olivo (foto mia); (a destra) le ramificazioni dell ife fungine (fonte: Weil & Brady, The Nature and Properties of Soils, 15th Edition, Pearson).
Scendendo a livello cellulare, le radici possiedono i peli radicali (ramificazioni più sottili nella foto qui in basso, piantine di Arabidopsis thaliana; scala: 0.5 mm; Sofo et al., Physiologia Plantarum 149: 487–498. 2013), che formano estroflessioni del rizoderma. Quindi possiamo definire le radici come una specie di intestino estroflesso, dove i villi sono le radici laterali e i microvilli i peli radicali. Tra peli radicali e ramificazioni laterali, la radice di una piantina minuscola di Arabidopsis, dal peso fresco di pochi milligrammi, può arrivare a 1 cm2 di superficie, 20 cm di lunghezza totale e 60-70 radici laterali (Sofo et al., Journal of Plant Physiology 216: 174-180).
Grazie a questa elevatissima superficie specifica, le radici riescono a svolgere la loro funzione assorbenti anche nelle condizioni più proibitive in suoli poverissimi e impervi, come nei due casi qui in basso.

Pino d’Aleppo su costone calcareo, spiaggia di Vignanotica, Gargano (FG) (foto mia).

Le radici contorte e fortemente lignificate del timo selvatico (Thymus vulgaris), Murgia Timone (MT) (foto mia).
Analogamente, le foglie e le chiome delle piante non sono disposte casualmente, ma tendono a intercettare al meglio la luce, a perdere meno acqua possibile e ad assorbire più CO2. Ad esempio, tra chiome di alberi della stessa specie o di specie diverse in una foresta c’è competizione per la luce. Anche in una stessa pianta, le foglie si dispongono secondo un preciso schema (fillotassi) per nulla casuale, e lo stesso fanno i rami. Vi mostro due figure che rendono l’idea, ma questa è una storia di ci parleremo più in là.

Le ramificazioni di una pianta rampicante di vite canadese (Ampelopsis brevipedunculata) (foto mia). Disposizione casuale o causale?

Faggi nel Parco Nazionale del Pollino (foto mia). Qui è visibile l’ “immersione” degli alberi nell’atmosfera.




