Darwin e i suoi erbari
Rivelare le storie inedite dietro alle specie di Darwin che hanno plasmato l’evoluzione
Il Progetto di Digitalizzazione di Kew sta stimolando un nuovo studio sulle collezioni di piante di Charles Darwin provenienti dal suo famoso viaggio a bordo dell’HMS Beagle.
Quasi due secoli dopo che il giovane Charles Darwin, all’età di 22 anni, partì per il suo viaggio intorno al mondo a bordo dell’HMS Beagle – un viaggio che ispirò la sua visione dell’evoluzione – le specie vegetali che raccolse continuano a fornire nuovi spunti e rimangono un’area attiva di ricerca curatoriale. L’Erbario di Kew, uno dei più grandi al mondo, è un importante centro delle collezioni di piante di Darwin. Dei 1.400 campioni vegetali raccolti da Darwin a bordo dell’HMS Beagle, circa 450 sono conservati a Kew, mentre la maggior parte situata presso l’Erbario dell’Università di Cambridge.
Cosa contiene la collezione di KEW di campioni di Darwin?
La collezione di Kew rappresenta 75 famiglie di piante, raccolte da Darwin in luoghi come le isole Galápagos, la Terra del Fuoco e l’arcipelago di Chonos in Cile. Ma poiché nuove informazioni su chi abbia raccolto quale pianta emergono continuamente con la digitalizzazione e la catalogazione digitale dei campioni, resi disponibili online tramite il progetto di digitalizzazione, nessuno conosce con certezza l’intero elenco dei campioni di Darwin.
Il compito di identificare ogni singola pianta raccolta da Darwin, o da qualsiasi collezionista dell’era vittoriana, è reso più difficile dal fatto che all’epoca era pratica comune attaccare più campioni vegetali su un unico foglio di erbario. Ciò crea problemi poiché i fogli devono essere archiviati in base a uno solo dei taxa presenti, quindi qualsiasi materiale attaccato a un campione non correlato è molto difficile da rintracciare. Ora un ambizioso progetto per digitalizzare l’intera collezione di più di 8 milioni di campioni di piante e funghi di Kew sta mettendo in luce alcuni angoli nascosti di questo unico archivio botanico.
Riscoperta di campioni storici
Nel processo, gli scienziati stanno scoprendo campioni di collezionisti precedenti che sono rimasti inosservati per anni. Se ciò includerà tesori sconosciuti di Darwin resta da vedere. Ciò significa che mentre il viaggio del Beagle è entrato da tempo nei libri di storia, le collezioni di piante di Darwin sono soggette a continue indagini e a una certa incertezza. Ciò che non è in discussione, tuttavia, è l’importanza storica e botanica dei campioni che ha raccolto mentre il Beagle viaggiava attraverso l’Atlantico, attorno all’America del Sud e ritorno in Inghilterra attraverso il Pacifico nel suo viaggio di esplorazione per l’Ammiragliato britannico dal 1831 al 1836.
Come Darwin giunse alla teoria della selezione naturale
Di particolare rilevanza sono il gran numero di campioni delle isole Galápagos, dove Darwin scoprì che le specie di piante e animali differivano da un’isola all’altra, anche se molte delle isole non erano lontane tra loro. Fu mentre rifletteva su questo che Darwin iniziò a formulare le sue idee rivoluzionarie sull’evoluzione della vita sulla Terra e sull’idea che una specie potesse generare più specie, ognuna sfruttando nicchie diverse.
E’ noto che Darwin fu ispirato dall’evoluzione adattativa dei fringuelli delle Galápagos, che hanno becchi diversi da un’isola all’altra, ma le piante giocarono anche un ruolo cruciale nello sviluppo della sua teoria sull’origine delle specie. Nel processo di raccolta di una vasta gamma di specie vegetali, creò una preziosa risorsa botanica che rimane fondamentale per la comprensione moderna dell’ecologia fragile dell’area. Prima della visita di Darwin nel 1835, si sapeva molto poco sulla gamma di fauna delle isole Galápagos e la sua visita fu trasformativa. Dopo cinque settimane di duro lavoro, i suoi sforzi di raccolta avevano coperto circa il 24% di tutta la flora delle isole conosciuta oggi.
Il ruolo delle piante nella teoria di Darwin
Catalogare e comprendere la rilevanza di ciò che aveva trovato fu un processo lungo e avrebbe richiesto un’altra decade per essere portato a termine. Darwin imballò con cura e inviò a casa campioni dal Beagle al suo mentore, il professor John Henslow a Cambridge, e i due uomini scambiarono lettere in cui Darwin esponeva le sue teorie in via di sviluppo. Gli Archivi di Kew contengono 44 di queste lettere. Le piante emersero successivamente come avendo un ruolo fondamentale nello sviluppo della teoria di Darwin sulla selezione naturale. I metodi innovativi di insegnamento di Henslow, tra cui l’uso di collezioni erbarie e conferenze illustrate, espose Darwin agli intricati schemi di variazione all’interno delle specie vegetali.
Le collezioni di piante di Darwin dal suo viaggio trasformativo, inviate a Henslow, gettarono le basi per la sua comprensione delle popolazioni e della variazione all’interno di esse. Queste prime esplorazioni botaniche, guidate dalla tutoria di Henslow, prefigurarono sottilmente i concetti evolutivi che avrebbero successivamente rivoluzionato la biologia. Fu solo nel 1843 che il giovane Joseph Dalton Hooker, figlio del direttore dell’epoca dei Giardini di Kew, accettò di esaminare dettagliatamente le piante delle Galápagos di Darwin.
Charles Darwin e Joseph Hooker discutono le scoperte di Darwin
Hooker presentò i suoi risultati a tre riunioni della Linnean Society nel 1845, descrivendo decine di nuove specie e concludendo che più della metà delle piante delle Galápagos non si trovava da nessun’altra parte nel mondo. In altre parole, erano endemiche e, in molti casi, limitate a singole isole. Hooker e Darwin, che divennero amici stretti, si impegnarono in una vivace corrispondenza sulle piante delle Galápagos mentre Darwin perfezionava la sua teoria dell’evoluzione.
In una famosa lettera del 1844 a Hooker, Darwin confidò per la prima volta la sua convinzione che le specie non fossero immutabili, aggiungendo che dire una cosa del genere era “come confessare un omicidio”. Di particolare interesse per Darwin e Hooker erano le piante della famiglia delle Asteraceae a causa del grande numero di generi e specie endemiche in questo gruppo. Diverse di queste piante sono presenti in modo prominente nell’Erbario di Kew, e la famiglia botanica è anche al centro degli sforzi moderni per proteggere la delicata flora delle Galápagos.
Gli ambientalisti sono preoccupati per il progressivo impoverimento delle foreste delle Galápagos di Scalesia, un genere di Asteraceae che forma alberi giganti con fiori a forma di margherita. Le piante sono uniche nelle Galápagos e ciascuna delle specie mostra una notevole adattamento alle condizioni di crescita nelle diverse isole. Sfortunatamente, ora sono minacciate dalla deforestazione, dai cambiamenti climatici e dalla diffusione delle piante di more invasive, che impediscono alla prossima generazione di alberi di crescere.
Accelerare la scienza utilizzando i campioni di Darwin
Oggi, un obiettivo chiave della Charles Darwin Foundation nelle Galápagos è salvare la specie Scalesia cordata, un albero endemico della parte meridionale dell’isola di Isabela che è sull’orlo dell’estinzione. Il progetto di digitalizzazione a Kew è ancora nelle fasi iniziali, con altri sei milioni di campioni da digitalizzare entro il 2026, il che significa che un tesoro di campioni di piante e i loro segreti devono ancora essere scoperti. Le collezioni di Kew sono fondamentali sia per la ricerca attuale che per le generazioni future per affrontare le nostre sfide più grandi, compresi i cambiamenti climatici e la perdita di biodiversità.

































 Sequoie giganti presso il Sequoia National Park (Ph: A. Sofo).
Sequoie giganti presso il Sequoia National Park (Ph: A. Sofo).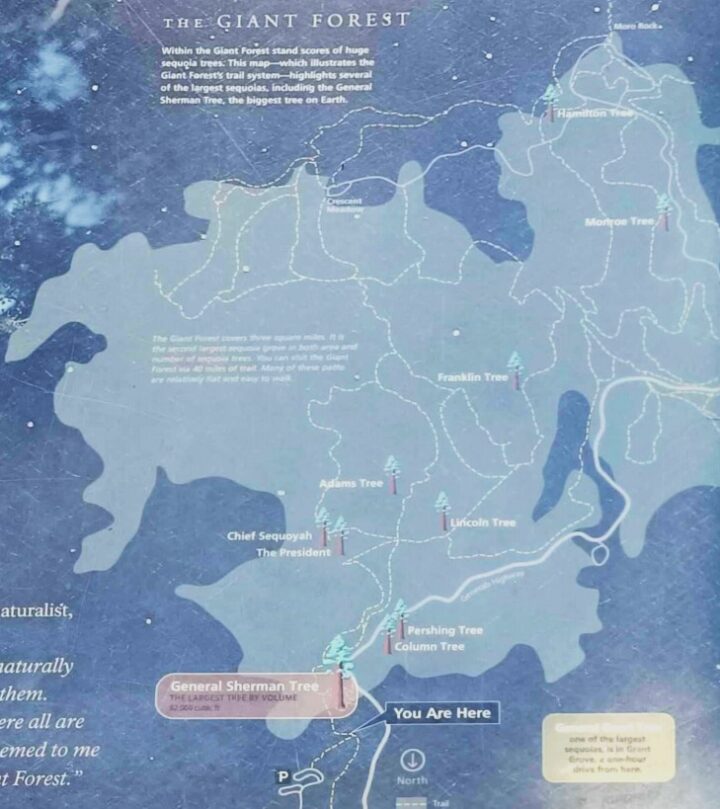



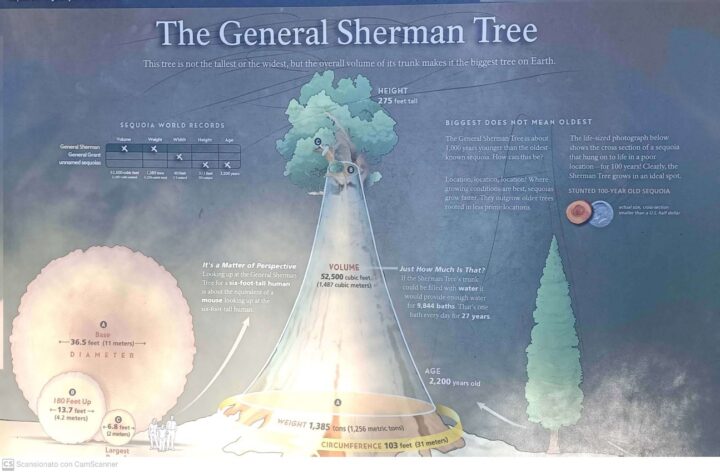

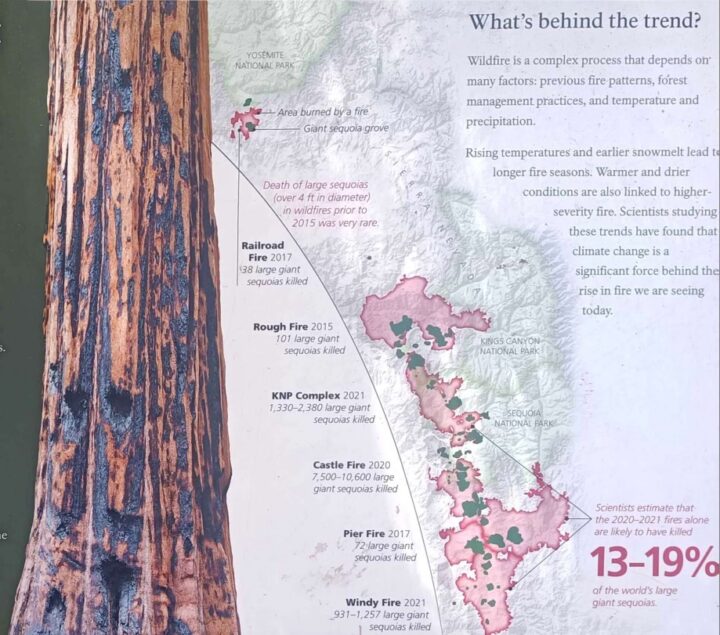
 Sezione trasversale di una sequoia gigante.
Sezione trasversale di una sequoia gigante.









