La gioia di contenere cloroplasti

“Bene, Caporale Westerburg,” – disse di nuovo il Dottor Harris – “Perché pensa di essere una pianta?”
Il Caporale guardò in alto timidamente. Si schiarì la voce: “Signore, io sono una pianta, non solo lo penso. E’ da parecchi giorni ormai che sono una pianta”.
“Lo vedo.” –annuì il Dottore – “Vuole dire che non è sempre stato una pianta?”
“No, signore. Sono diventato una pianta solo recentemente.”
“E cos’era prima che diventasse una pianta?”
“Bene, signore, ero come voi tutti.”
(da “I pifferai”; Philip K. Dick)
Con le sue tipiche ambientazioni post apocalittiche e tinte fosche, Dick fa raccontare al Caporale Westerburg di essere una pianta. E difatti si comporta da pianta, smettendo di lavorare. Non solo; anche i suoi colleghi, di scorta su un asteroide al confine del Sistema Solare, sono affetti dalla stessa psicosi: sono convinti di essere diventati piante e passano tutta la loro giornata seduti al sole, attribuendo il motivo del loro cambiamento a misteriosi “pifferai”, una tribù indigena che vivrebbe sull’asteroide. Il Dottor Harris, che li ha in cura, si reca sull’asteroide e capisce che la psicosi deriva solamente dall’inconscio desiderio di non avere più alcuna responsabilità e abbracciare uno stile di vita semplice come quello degli alieni che vivono sull’asteroide.
Gli omini verdi sono una caratteristica tipica dei film di fantascienza e dei supereroi (buoni o cattivi che siano (ricordate Hulk, Lizard, i capelli Joker?). Già in “Sotto le lune di Marte” (1912), Edgar Rice Burroughs descrisse varie specie di marziani, tra cui una con la pelle verde. Tra il 1920 e il 1950 gli omini verdi diventarono popolari grazie alle riviste di fantascienza con le storie di Flash Gordon e Buck Rogers. A me vengono invece in mente gli elfi del bosco e le crature verdi a tre occhi di Toy Story, ma sono un caso a parte.
Pelle e capelli verdi sono sempre stati associati a individui minacciosi, proprio perché è un colore tipico delle piante e inusuale negli animali con cui l’uomo ha confidenza. Molti rettili hanno livree che colorate piò o meno di verde e spesso sono considerati animali malvagi o infidi, perlomeno pericolosi.
Ma perché gli esseri umani non sono verdi? E perché non fanno fotosintesi come le piante? In fin dei conti, ci risparmieremmo un bel po’ di fastidio e sarebbe comodo essere autotrofi.
Le piante sono verdi perché molte delle loro cellule contengono cloroplasti, organelli intracellulari in cui avviene la fotosintesi. I cloroplasti contengono clorofilla, il principale pigmento dal tipico colore verde che cattura l’energia della luce solare (in particolare quella rossa e blu) e dà avvio alla fotosintesi, vitale per la pianta. La sua atruttura chimica è curiosamente affine a quella dell’emoglobina, con una differenza principale: il magnesio, che nella clorofilla sostituisce il ferro dell’emoglobina (ne avevamo parlato in un post di qualche tempo fa).
I cloroplasti hanno una storia evolutiva piuttosto interessante, perché originariamente erano cianobatteri a vita libera, indipendenti dalle piante. I cianobatteri sono famosi perché sono batteri – ma non gli unici – che hanno “inventato” la fotosintesi, un processo che sfrutta l’energia solare per fissare il carbonio atmosferico gassoso (CO2) in carbono solido (zuccheri), utilizzando acqua come donatore di elettroni. L’invenzione ebbe, per così dire, molto successo e di conseguenza si diffuse velocemente. Anche altri gruppi batterici, più o meno indipendentemente e per vie leggermente diverse, arrivarono alla fotosintesi.
In un momento di grande ispirazione scientifica, Lynn Margulis si rese conto che i cloroplasti all’interno delle cellule vegetali erano cianobatteri “addomesticati” e catturati centinaia di milioni di anni fa da cellule eucariote (fornite di nucleo) ed eterotrofe (dovevano assimilare carbonio organicato), che quindi non facevano fotosintesi: nacque così la cellula vegetale e le prime alghe unicellulari.
Posso immaginare – e ci sarà sicuramente stato – il momento preciso in cui un antenato unicellulare della piante inghiottì un cianobatterio e, invece di digerirlo, si rese conto che si trattava di un utile parente acquisito. Grazie a quel momento, noi animali siamo qui oggi. Ancora più fondamentale per il funzionamento di tutti gli organismi superiori è un secondo organello, chiamato mitocondrio. Margulis comprese che anche questo era stato una volta un batterio vivente libero che sfruttava l’energia chimica rinchiusa in substrati zuccherini, come il glucosio. Quindi le cellule di piante sono praticamente chimere, contenenti DNA dell’ospite originale più quello dei due batteri catturati. Questa teoria è nota come teoria endosimbiontica.
Contenere cloroplasti porta un vantaggio enorme e immediato. Le cellule animali hanno solo i mitocondri, che permettono loro di ossidare il glucosio e sfruttare l’energia chimica risultante per il loro metabolismo, ma devono trovare una fonte di carbonio esterna nel glucosio (sono appunto eterotrofi). Questo significa dedicare una parte consistente del loro tempo per localizzare, appropriarsi e consumare del cibo. Le piante, d’altra parte, non devono preoccuparsi (sono autotrofe: la loro fonte di carbonio è inorganica). Esse possono utilizzare i cloroplasti per produrre il proprio glucosio, che possono poi passare ai mitocondri per liberare l’energia chimica, quando è necessario.
Se le piante non si curano di trovare glucosio, gli animali purtroppo non possono fare la stessa cosa. In realtà, alcuni animali hanno adottato un’altra strategia e si sono spinti oltre. Il cloroplasto era semplicemente un’invenzione troppo geniale e molti altri organismi cercarono quindi di mendicare, prendere in prestito o rubare cloroplasti altrui, principalmente da alghe unicellulari a vita libera. Questo processo è noto come endosimbiosi secondaria, per distinguerlo dall’endosimbiosi primaria, come quella prima descritta a riguardo dell’antenato delle piante che aveva inghiottito un cianobatterio a vita libera.
Non è del tutto chiaro il motivo per cui l’endosimbiosi secondaria sembra essersi verificata molte volte, mentre quella primaria si sia verificata una sola volta (anche se recentemente gli scienziati hanno scoperto un secondo esempio di un’endosimbiosi primaria in una specie di ameba (Paulinella), la quale sembra aver “addomesticato” un cianobatterio, riproponendo così l’evento che diede origine alle piante terrestri). Il trasferimento del cloroplasto mediante endosimbiosi secondaria ha dato vita a tutta una serie di organismi ecologicamente importanti, la maggior parte dei quali unicellulari. Questi organismi, tra i quali ricordiamo diatomee, dinoflagellati e euglenoidee (i primi due gruppi alghe e le ultime protisti fotosintetici) derivano da acquisizioni indipendenti di cloroplasti appartenenti precedentemente ad alghe. Euglena viridis, forse il più famoso rappresentante delle euglenoidee, è una specie fotosintetica come un’alga unicellulare ma può anche cambiare il suo metabolismo in eterotrofo quando la luce non c’è. E’ quindi un organismo mixotrofo e per questo non è considerato un’alga ma un protozoo, sebbene fotosintetico (nella foto qui in basso, una raffigurazione di Euglena viridis in computer grafica).
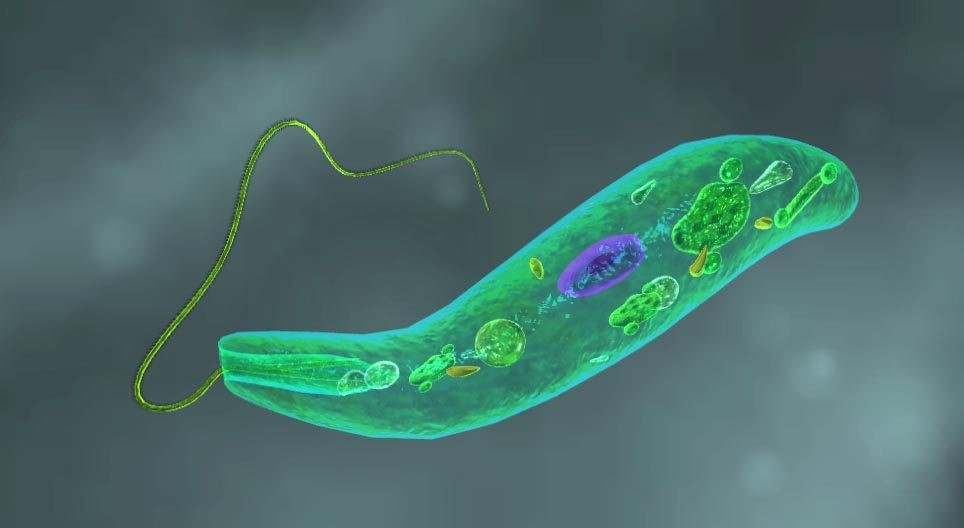
Gli organismi unicellulari fotosintetici sono a loro volta stati catturati da animali pluricellulari. Queste simbiosi si sono evolute in maniera indipendente molte volte e il rapporto tra l’ospite e il fotosimbionte ha assunto varie forme. Ad esempio, ci sono lumache marine verdi (Elysia), che rubano cloroplasti dalle alghe su cui pascolano (foto in basso). Queste non possono mantenere cloroplasti per lungo tempo e quindi hanno bisogno di un approvvigionamento costante. C’è ancora un dibattito molto acceso sul fatto che i cloroplasti siano davvero indispensabili per la lumaca.

D’altra parte, molti organismi marini, come coralli, molluschi giganti e ascidie, sono totalmente dipendenti dai loro simbionti e non posono vivere senza di loro. La fotosintesi è ospitata anche nella salamandra “solare” Ambystoma maculatum (video qui in basso).
C’è poi il caso della Vespa orientalis, che possiede dei veri e propri pannelli fotovoltaici nell’addome e che, grazie alla xantopterina (il principale pigmento responsabile del colore giallo delle bande addominali, che si alternano alle bande nere, contenenti invece melanina) non solo assorbe energia solare e la dissipa come calore, ma addirittura è in grado di trasformarla in potenziale elettrochimico, con produzione di ATP (la principale molecola che accumula energia). In parole povere, il processo ricorda molto la fotosintesi, in cui la clorofilla converte radiazione solare prima in potenziale elettronico e poi in potenziale chimico sotto forma di ATP e NADPH.
Quindi, se lumache e ascidie possono trarre vantaggio dalla fotosintesi, perché noi non possiamo? Il tutto sta nel considerare il bilancio energetico di un grande animale multicellulare e molto attivo, quale un essere umano. Ogni giorno un adulto umano richiede l’equivalente del proprio peso corporeo in ATP, una molecola che immagazzina l’energia chimica rilasciata dall’ossidazione del glucosio. Per produrre circa 60 kg di ATP, un uomo adulto di media stazza richiede circa 700 g di glucosio al giorno. Considerando i tassi massimi noti per la fotosintesi nelle piante superiori, e che la superficie della pelle di un adulto è di circa 1,6 m2, un uomo con la pelle verde sarebbe capace di produrre soltanto un deludente 1% della sua domanda giornaliera di glucosio mediante fotosintesi. Quindi, per soddisfare le sue richieste di energia, l’ipotetico uomo fotosintetico dovrebbe avere una superficie di pelle equivalente a quella di un campo da tennis. Eppure, esempi di fotosintesi artificiale non mancano e recentemente è stato trasformato un batterio (Moorella thermoacetica) non fotosintetico al fine di indurlo a fotosintetizzare e a trasformare energia solare e anidride carbonica in acido acetico. Si tratta “solo” di batteri e non di esseri umani, ma è un notevole passo in avanti.
Le piante risolvono questo problema perché, a parità di volume, hanno una maggiore superficie esterna fotosintetica (epidermide delle foglie e degli altri organi verdi). Inoltre, l’epidermide delle foglie è più trasparente alla luce e le foglie si fanno attraversare facilmente dalla radiazione solare, considerando il loro piccolo spessore. In più, a livello cellulare, i cloroplasti contengono sacculi impilati e contorti, che nell’insieme hanno un’ampia superficie totale, chiamati tilacoidi, nelle cui pareti le molecole di clorofilla si infilzano come chiodi. E, per finire, i cloroplasti si possono muovere all’interno delle cellule, disponendosi secondo posizioni ottimali, al fine di catturare più luce possibile.
Dobbiamo quindi concludere, a malincuore, o che gli alieni verdi dovrebbero essere sostanzialmente più strani di quanto essi non siano attualmente ritratti nei film e nei fumetti, oppure che la fotosintesi che si è evoluta su altri pianeti, colonizzati da piccoli omini verdi, sia seriamente più efficiente di quella terrestre.
Qui, sulla triste Terra, non ci resta che rimanere eterotrofi, completamente dipendenti dalle piante per i carboidrati di cui abbiamo bisogno, e consumatori diretti di clorofilla solo per le sue preziose virtù extranutrizionali.









